



Introduction
Sweet pepper (Capsicum annuum), also known as bell pepper, is an annual, subshrub or shrub belonging to the family Solanaceae and grows primarily in the seasonally dry tropical biome (Plants of the World Online, 2024). Sweet pepper fruits are consumed worldwide as a popular vegetable. In Korea, sweet peppers are mostly grown in greenhouses, while chili peppers are mainly grown in fields.
Over 50 fungi have been recorded to cause pepper diseases in USA (Farr et al., 1989). In Korea, about 20 fungal diseases have been reported to occur in chili pepper, and the major diseases that occur on chili pepper fruits are anthracnose, black mold, gray mold, black dot fruit rot, and internal fruit rot (Korean Society of Plant Pathology, 2024). We observed black mold symptoms on fruits of sweet pepper plants grown in a greenhouse located in Suwon, Korea, during the harvest season in 2022 and 2023. We examined morphological characteristics of fungal isolates from the diseased fruits and found that they belonged to the genus Alternaria (Ellis, 1971). Simmons (2007) summarized 275 Alternaria spp. identified based on morphological characteristics. However, many Alternaria spp. have been known to be very difficult to distinguish by their morphological characteristics alone. Therefore, taxonomy of Alternaria spp. has been studied based on DNA sequence data in combination with morphology (Woudenberg et al., 2013; Woudenberg et al., 2015).
Alternaria spp. have been reported to cause various diseases in plants (Ellis, 1971; Rotem, 1994). They cause diseases based on the specificity to the family, genus or species of the host plants, but some Alternaria spp. are known to be unable to cause diseases due to non-pathogenic strains or variety resistance of the host plant (Rotem, 1994). This study was conducted to identify the unknown Alternaria sp. isolates from the diseased fruits of sweet pepper based on morphological characteristics and molecular phylogenetic analyses. In addition, pathogenicity of the isolates was tested to sweet pepper fruits to determine the pathogen causing black mold in sweet pepper.
Materials and Methods
Disease survey and isolation of fungi.
In July 2022 and 2023, we surveyed occurrence of black mold on fruits of sweet pepper plants grown in a greenhouse located in Suwon, Korea. During the disease survey, three sites were observed in the greenhouse, and 100 fruits at each site were investigated for the disease incidence. The diseased fruits were collected from the investigated greenhouse for isolation of pathogens. A conidial suspension was prepared from conidial masses on the fruits using sterile distilled water and streaked on 2% water agar (WA) plates using a sterile loop. The WA plates were incubated at 25°C for 24 hr, and germinated conidia were picked up under a dissecting microscope (Nikon SMZ 1780; Nikon, Tokyo, Japan) and transferred to new WA plates. Single-conidium isolates were obtained from the WA plate cultures after 5 days of incubation at 25°C. The isolates were cultured on potato dextrose agar (PDA) slants for identification and pathogenicity tests.
Investigation of morphological characteristics.
To investigate morphological characteristics of the isolates, each isolate was incubated on PDA at 25°C in the dark for 15 days. After incubation, the colony morphology of the isolates was observed, and the shape and size of 30 conidiophores and conidia of each isolate formed on the medium were investigated using a compound microscope (Nikon Eclipse Ci-L; Nikon). In addition, a mycelial disk of 6 mm from each isolate grown on PDA was transferred to WA and incubated at 25°C in the dark for 10 days to examine formation patterns of conidia on conidiophores using the compound microscope.
DNA sequencing and phylogenetic analysis.
Genomic DNA of the isolates was extracted using the protocol in a previous study (Dong et al., 2022), with slight modifications. In polymerase chain reaction (PCR) experiments, glyceralde-hyde-3-phosphate dehydrogenase (GAPDH), alternaria major allergen (Alt a 1), RNA polymerase II second largest subunit (RPB2), and translation elongation factor 1-alpha (TEF1) gene regions were amplified with gpd1 and gpd2 (Berbee et al., 1999) for GAPDH, Alt-for and Alt-rev (Hong et al., 2005) for Alt a 1, RPB2-5F2 (Sung et al., 2007) and fRPB2-7cR (Liu et al., 1999) for RPB2, and EF1-728F and EF1-986R (Carbone and Kohn, 1999) for TEF1. Conditions of PCR amplification for all the genes were followed as described in the previous studies (Woudenberg et al., 2013; Woudenberg et al., 2014). DNA Free-Multiplex Master Mix (Cellsafe, Yongin, Korea) was used to prepare PCR products of the isolates following the manufacturer's instructions. Purification for the PCR products was done using the Universal DNA Purification Kit (Tiangen, Beijing, China) following the manufacturer's protocol. Sequencing for the PCR products was conducted at Bionics Co., Ltd. (Seoul, Korea) with the same primers. The sequences were adjusted by SeqMan II (DNASTAR Inc., Madison, WI, USA) if necessary. The sequence data were deposited in the National Center for Biotechnology Information (NCBI) Genbank.
The sequences of the isolates and the relevant sequences of Alternaria spp. from the previous study (Woudenberg et al., 2015) were aligned together using MUSCLE (Edgar, 2004). Alternaria solani (CBS 109157) was used as an outgroup tax-on. The multiple sequence alignments were processed and improved, if necessary, with MEGA version 7 software (Kumar et al., 2016). The alignments were concatenated to construct a phylogenetic tree using the neighbor-joining method with a maximum composite likelihood model performing 1,000 bootstrap replicates by MEGA version 7 software (Kumar et al., 2016). Bootstrap values were shown at the nodes of the phylogenetic tree.
Pathogenicity test.
The isolates from sweet pepper fruits were used for pathogenicity tests to sweet pepper fruits of two varieties (SP-504 and SP-505). To make an inoculum, each isolate was transferred to PDA and incubated at 25°C in the dark for 15 days, and then conidia formed on the medium were suspended in sterile distilled water to prepare a conidial suspension. The concentration of each conidial suspension was adjusted to 3−4×105 conidia/ml using a hemocytometer.
The sweet pepper fruits of each variety were immersed in 1% sodium hypochlorite (NaOCl) solution for 5 min for surface sterilization and then rinsed with sterile distilled water. After removing moisture, the fruits were placed in humid plastic boxes (29.0×22.5×12.0 cm), and 30 μl of each conidial suspension was dropped on the middle part of the fruits for inoculation test. In the case of wound inoculation, the central portion of the conidial suspension droplet on the fruit was stabbed to a depth of about 1 mm using the tip of a needle. The same quantity of sterile distilled water was used to control sweet pepper fruits. The plastic boxes containing the inoculated sweet pepper fruits were incubated at 25°C for 10 days, and then the result of the inoculation tests was investigated. The inoculation test was performed in three repetitions.
Results and Discussion
Disease symptoms and incidence.
In July 2022 and 2023, we observed sporadic occurrence of black mold symptoms on fruits of sweet pepper plants grown in a greenhouse located in Suwon, Korea. In the early stages of the symptoms, lesions on the fruits were slightly dented in a soft state. As the disease progressed, the lesions expanded to a circular shape, and numerous black molds formed on the lesions (Fig. 1A-D). Severely diseased fruits became soft and rotten. The incidence of black mold on the fruits in the investigated greenhouse was 5−24% (average 14.8%) in variety SP-504 (yellow and elongate type) and 1−8% (average 5%) in variety SP-505 (red and round type) (Table 1).
Fig. 1.
Symptoms of black mold on sweet pepper fruits. Symptoms observed on the fruits of varieties SP-504 (A, B) and SP-505 (C, D) in the greenhouse. Symptoms on a non-wounded fruit (E) and a wounded fruit (F) of variety SP-504 induced by artificial inoculation with the isolate PRAL-10 of Alternaria alternata. Symptoms on a wounded fruit (H) of variety SP-504 induced by artificial inoculation with the isolate PRAL-10 of A. alternata. (G, I) Non-inoculated control fruits of varieties SP-504 and SP-505, respectively.

Table 1.
Occurrence of black mold on sweet pepper fruits of two varieties grown in a greenhouse located in Suwon, Korea in July 2022 and 2023
| Variety of sweet pepper | Fruit type | Diseased fruits a (%) |
|---|---|---|
| SP-504 | Yellow and elongate | 5−24 (average 14.8) |
| SP-505 | Red and round | 1−8 (average 5.0) |
Morphological identification of isolates.
We obtained four single-conidium isolates of fungi from the lesions on sweet pepper fruits and examined morphological characteristics of the isolates. Conidia were obclavate, ovoid or ellipsoidal, pale to dark brown, with 1−7 transverse septa, 1−2 or no longitudinal septa (Fig. 2A), and measured 12.3−65.3×8.2−17.5 μm (average 35.5×11.6 μm) (Table 2). Conidia formed in long chains on conidiophores (Fig. 2B), and the colony of the isolates cultured on PDA displayed gray to black (Fig. 2C). All the isolates were identified as Alternaria alternata based on their morphological characteristics according to the previous descriptions (Ellis, 1971; Simmons, 2007).
Fig. 2.
Morphology and cultural appearance of Alternaria sp. isolated from sweet pepper fruits. (A) Conidia isolated from a lesion of black mold. (B) A conidial chain produced on a conidiophore in WA culture. (C) A colony of an isolate grown on potato dextrose agar at 25°C in the dark for 15 days.

Table 2.
Morphological characteristics of Alternaria alternata isolates from diseased sweet pepper fruits and the fungal morphology described by previous researchers
| Structure | Present isolates | Ellis (1971) | Simmons (2007) |
|---|---|---|---|
| Conidiophores | Simple or branched, straight or flexuous, sometimes geniculate, 8.2−103.2×3.5−5.5 μm (average 37.5×4.3 μm) | Simple or branched, straight or flexuous, sometimes geniculate, up to 50 μm long, 3−6 μm thick | Simple or 1−3 branched, geniculate, comparatively short, 40−70×3−4 μm |
| Conidia | Formed in long chains, obclavate, ovoid or ellipsoidal, pale to dark brown, with 1−7 transverse septa, 1−2 or no longitudinal septa, 12.3−65.3×8.2−17.5 μm (average 35.5×11.6 μm) on potato dextrose agar | Formed in long chains, obclavate, obpyriform, ovoid or ellipsoidal, pale to mid golden brown, with up to 8 transverse septa, several longitudinal septa, 20−63×9−18 μm (average 37×13 μm) | Formed in chains, ovoid, ellipsoid or subsphaeroid, olivaceous, dull grey-green-brown, with 1−7 transverse septa and 1−2 or no longitudinal septa, 13−43×8−14 μm on V8 juice agar |
| Beak of conidia | Short conical or cylindrical, pale brown, 2.0−17.8×2.3−5.0 μm (average 6.5×3.5 μm) | Short conical or cylindrical, pale, not more than one third the length of the conidium, 2−5 μm thick | No description |
Molecular phylogeny.
The concatenated alignment of 11 ingroup taxa contained a total of 2,109 characters (564, 473, 736, and 336 characters for GAPDH, Alt a 1, RPB2, and TEF1 gene regions, respectively). Phylogenetic analysis based on concatenated sequence alignments of the GAPDH, Alt a 1, RPB2, and TEF1 gene regions revealed that the isolates clustered with three strains (CBS 121455, CBS 479.90, and CBS 102598) of A. alternata (Fig. 3). The A. alternata clade was distinguishable certainly with a significant bootstrap value from closely related Alternaria spp. such as Alternaria alstroemeriae and Alternaria gaisen. The sequence data of the genes obtained from the isolates were deposited in the NCBI Genbank under the designated accession numbers (Table 3).
Fig. 3.
Phylogenetic tree based on glyceraldehyde-3-phosphate dehydrogenase, alternaria major allergen, RNA polymerase II second largest subunit, and translation elongation factor 1-alpha gene sequence data for Alternaria alternata isolates from diseased sweet pepper fruits and reference species. Sequence data were obtained from the NCBI GenBank database. The phylogenetic tree was generated using the neighbor-joining method with a maximum composite likelihood model. The bootstrap values were provided at nodes. The scale bar represents the number of nucleotide substitutions per site. T, ex-type strain; R, representative strain.
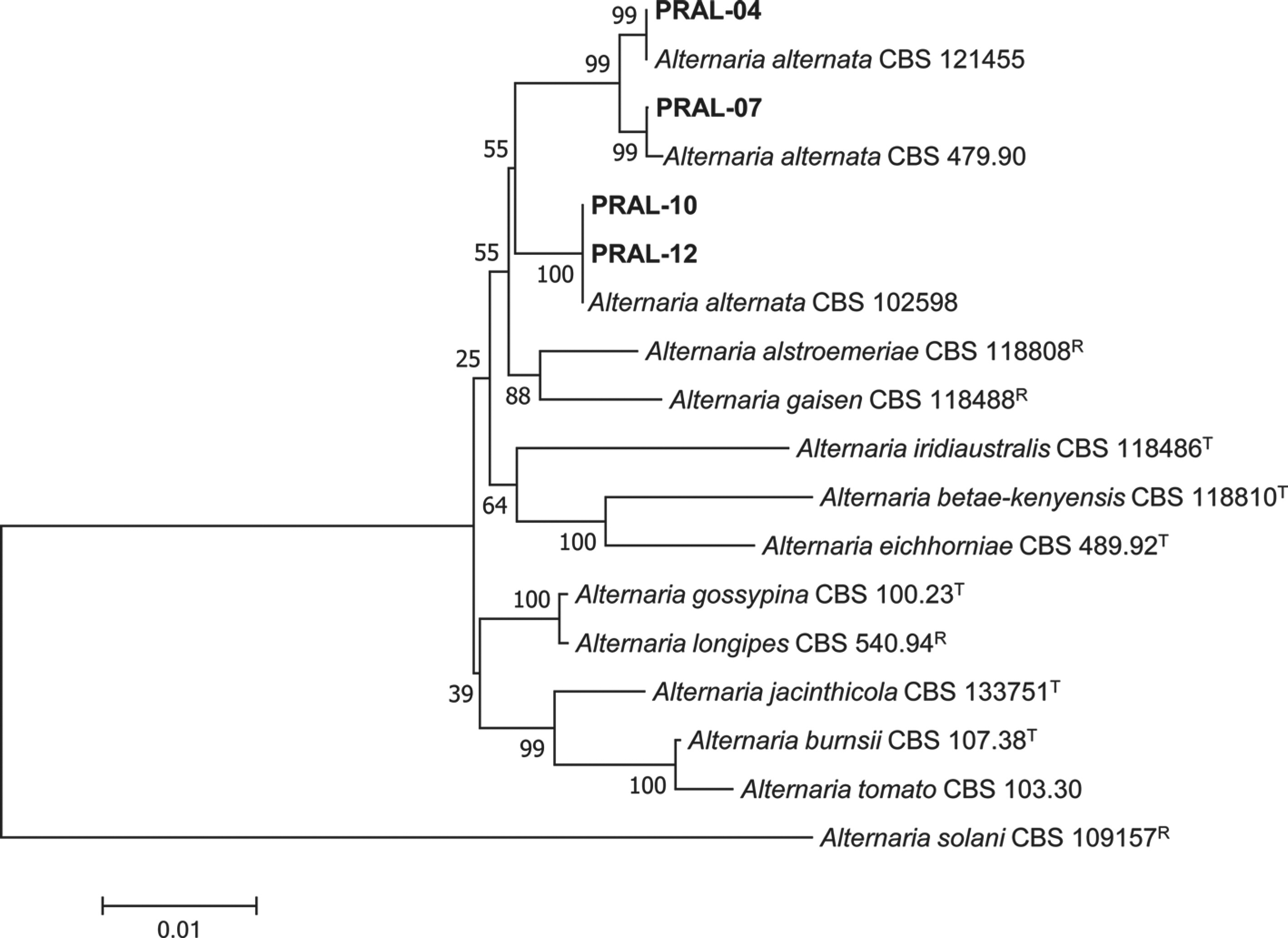
Table 3.
Isolates and strains of Alternaria spp. used for molecular phylogenetic analyses in this study
| Alternaria spp. | Isolate/strain a | Host | Locality | Genbank accession number | |||
|---|---|---|---|---|---|---|---|
| Alt a 1 | GAPDH | RPB2 | TEF1 | ||||
| A. alternata | PRAL-04 | Capsicum annuum | Korea | PP272035 | PP272039 | PP272043 | PP272047 |
| A. alternata | PRAL-07 | Capsicum annuum | Korea | PP272036 | PP272040 | PP272044 | PP272048 |
| A. alternata | PRAL-10 | Capsicum annuum | Korea | PP272037 | PP272041 | PP272045 | PP272049 |
| A. alternata | PRAL-12 | Capsicum annuum | Korea | PP272038 | PP272042 | PP272046 | PP272050 |
| A. alstroemeriae | CBS 118808 R | Alstroemeria sp. | USA | KP124153 | KP123845 | KP125071 | KP124764 |
| A. alternata (A. pellucida T) | CBS 479.90 | Citrus unshiu | Japan | KP124174 | KP123870 | KP125095 | KP124787 |
| A. alternata (A. broussonetiae T) | CBS 121455 | Broussonetia papyrifera | China | KP124220 | KP123916 | KP125146 | KP124838 |
| A. alternata (A. citriarbusti T) | CBS 102598 | Minneola tangelo | USA | KP124184 | KP123878 | KP125105 | KP124797 |
| A. betae-kenyensis | CBS 118810 T | Beta vulgaris var. cicla | Kenya | KP124270 | KP123966 | KP125197 | KP124888 |
| A. burnsii | CBS 107.38 T | Cuminum cyminum | India | JQ646305 | KP123967 | KP125198 | KP124889 |
| A. eichhorniae | CBS 489.92 T | Eichhornia crassipes | India | KP124276 | KP123973 | KP125204 | KP124895 |
| A. gaisen | CBS 118488 R | Pyrus pyrifolia | Japan | KP124278 | KP123975 | KP125206 | KP124897 |
| A. gossypina | CBS 100.23 T | Malus domestica | Unknown | KP124280 | KP123977 | KP125208 | KP124899 |
| A. iridiaustralis | CBS 118486 T | Iris sp. | Australia | KP124284 | KP123981 | KP125214 | KP124905 |
| A. jacinthicola | CBS 133751 T | Eichhornia crassipes | Mali | KP124287 | KP123984 | KP125217 | KP124908 |
| A. longipes | CBS 540.94 R | Nicotiana tabacum | USA | AY278811 | AY563304 | KC584667 | KC584409 |
| A. tomato | CBS 103.30 | Solanum lycopersicum | Unknown | KP124294 | KP123991 | KP125224 | KP124915 |
| A. solni (Porri section) | CBS 109157 | Solanum tuberosum | USA | KC584139 | KJ718746 | KJ718585 | KJ718413 |
Pathogenicity.
The pathogenicity tests revealed that the isolates of A. alternata caused black mold lesions on sweet pepper fruits of two varieties, and there was a slight difference in the degree of lesion formation between the two varieties (Table 4). The isolates mostly induced large lesions on fruits of the two varieties in the wound inoculation, but only the isolates PRAL-10 and PRAL-12 small lesions on fruits of the variety SP-504 in the non-wound inoculation. No lesions formed on fruits of the variety SP-505 in the non-wound inoculation. The symptoms on the fruits induced by pathogenicity tests with the isolates were similar to those observed on fruits from the greenhouse investigated (Fig. 1E, F, H). No symptoms were observed on the control fruits (Fig. 1G, I). The inoculated isolates were re-isolated from the lesions.
Table 4.
Result of pathogenicity tests of Alternaria alternata isolates to sweet pepper fruits of two varieties
| Isolate No. | Lesion formation of isolates on fruits of sweet pepper varieties a | |||
|---|---|---|---|---|
| SP-504 (yellow and elongate type) | SP-505 (red and round type) | |||
| Non-wounded | Wounded | Non-wounded | Wounded | |
| PRAL-04 | − | ++ | − | ++ |
| PRAL-07 | − | ++ | − | + |
| PRAL-10 | + | ++ | − | ++ |
| PRAL-12 | + | ++ | − | ++ |
| Control | − | − | − | − |
A. alternata is known to cause various diseases in many plants and also a secondary infectious fungus that cannot cause disease in some plants (Ellis, 1971; Rotem, 1994). In addition, several strains of the fungus produce host-specific toxins and are highly pathogenic to a limited number of cultivars (Rotem, 1994). In Korea, it has been reported that black mold caused by A. alternata occurs on fruits of chili pepper (Kim and Yu, 1985) and tomato (Kim et al., 2020). The occurrence of black mold on sweet pepper fruits is first reported in this study.
Alternaria rot of pepper fruits caused by A. alternata occurs at the site of wounds received during harvest, or on tissue that has been damaged by chilling, sunscald, calcium deficiency, insect injury, or heat (Bartz, 2003). Damage in tomato caused by black mold is generally limited to senescent or wounded fruit due to sunscald, insect injury, and blossom-end rot (Davis and Paulus, 2014). In the present study, pathogenicity tests revealed that black mold lesions were formed better on sweet pepper fruits by wound inoculation than by non-wound inoculation, as previously reported in a study of tomato black mold (Kim et al., 2020).
The disease survey in the present study showed that occurrence of black mold on sweet pepper fruits in the greenhouse was more severe in variety SP-504 than in variety SP-505. Pathogenicity tests also revealed that the lesion formation was better in variety SP-504 than in variety SP-505, suggesting that susceptibility of sweet pepper fruits to the disease differs between the varieties. It is considered that more studies on the variety resistance of sweet pepper fruits to black mold and control of the disease are needed in the future.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






