



Momordica charantia, commonly known as bitter melon or bitter gourd, was the most extensively cultivated among subtropical crops such as asparagus, banana, coffee, dragon fruit, feijoa, guava, hooker chives, mango, okra, papaya, passion fruit, and turmeric until 2018 (Jeong et al., 2020). In Korea's agricultural landscape, the area dedicated to M. charantia cultivation averaged approximately 84.2 ha between 2017 and 2019. Within the specific region of Chungcheongbuk-do, the cultivation area for M. charantia averaged around 5 ha from 2017 to 2019 (Jeong et al., 2020). M. charantia is susceptible to various fungal diseases such as anthracnose and soft rot (Kim et al., 2015; Kwon and Jee, 2005). Bacterial soft rot diseases have been documented in Japan (Kubo et al., 2009). However, to date, there have been no reported instances of bacterial diseases affecting M. charantia in Korea.
Soft rot is a highly destructive ailment that has a global impact on vegetables, affecting regions where fleshy storage tissues of vegetables and ornamentals are present (Appy et al., 2023; Wu et al., 2023). Among bacterial plant pathogens, there exists a group of pectinolytic organisms known as soft rot Pectobacteriaceae, which encompasses two genera: Pectobacterium and Dickeya (Adeolu et al., 2016). Pectobacterium, recognized as one of the top 10 economically significant plant pathogenic bacteria, demonstrates a wide range of host compatibility, leading to the development of soft rot symptoms in both live plants and harvested crops (Ma et al., 2007; Mansfield et al., 2012). The invasion of the vascular systems of vegetables, ornamentals, and fruit trees by Pectobacterium strains can result in crop losses at various stages, including field production, harvest, storage, and transportation, with significant economic consequences (Czajkowski et al., 2011; Koh et al., 2012; P├®rombelon, 2002).
The genus Pectobacterium is known for its production of enzymes, including cellulase, pectinase, and polygalacturonase, which actively degrade plant cell walls and membranes (Hugouvieux-Cotte-Pattat et al., 2014; Maung et al., 2022). Under favorable climatic conditions, infected tissue exhibits a characteristic water-soaked, slimy, and rotten appearance (Barras et al., 1994; Hugouvieux-Cotte-Pattat et al., 1996).
Within the same species, the genus Pectobacterium encompasses diverse strains with varying biochemical, physiological, and genetic characteristics (Duarte et al., 2004; Portier et al., 2019). The use of molecular techniques for differentiation and advanced diagnostic methods has led to the reclassification of strains with distinct biochemical and physiological traits (Nabhan et al., 2012; Portier et al., 2019). Currently, this genus comprises 20 species with validly published names and several other proposed taxa are awaiting their validation (Hong et al., 2023b).
P. brasiliense, a member of the genus Pectobacterium, is recognized for its exceptional virulence. It was first identified as an atypical strain of Erwinia carotovora causing severe blackleg disease in Brazilian potato plants by Duarte et al. (2004). Initially suggested as a new subspecies of P. carotovorum, it was later classified as a separate species (Nabhan et al., 2012; Portier et al., 2019). P. brasiliense has recently gained worldwide attention as a significant threat to global potato production (Ûztürk and Umar, 2022).
The presence of soft rot induced by P. brasiliense in Korea was initially documented in 2012 in paprika (Choi and Kim, 2013). Since then, P. brasiliense has been identified in various plant species in Korea, including cucumber, graft cactus, melon, and paprika (Choi and Kim, 2013; Hong et al., 2023a; Park et al., 2022, 2023).
In August 2021, cases of soft rot diseases were observed on M. charantia fruits in Goesan, Chungcheongbuk-do, Korea. The affected fruits displayed a distinct transition to yellowing, followed by a subsequent collapse, revealing sunken lesions on the fruit surface and exhibiting typical soft rot symptoms (Fig. 1A, B). Tissue samples from the affected M. charantia were extracted for analysis. To isolate the pathogens, the affected M. charantia fruits underwent a surface sterilization process lasting 90 sec, using a 1% hypochlorite solution, followed by thorough rinsing with sterile distilled water. After sterilization, a 100 ╬╝l aliquot of liquid suspension was applied to the surface of a CVP medium (H├®lias et al., 2012). Distinctive colonies developed within the CVP medium after a three-day incubation period at 28┬░C. These colonies were characterized by their white-gray color, circular morphology, smooth edges, and noticeable cavities, primarily due to their pectin metabolism capability. To ensure the purity of the cultures, successive streaking techniques were employed on nutrient agar (NA; Difco, Franklin Lakes, NJ, USA) medium. One representative bacterial isolate, designated as KNUB-09-21, was selected for further comprehensive analysis. This strain was stored in 30% glycerol stocks at ŌłÆ20┬░C.
Fig.┬Ā1.
Display of soft rot symptoms in Momordica charantia plants from Goesan, Chungcheongbuk-do, Korea. (A) The infected fruits show a clear shift to yellow. (B) The area with the collapsed structure was softened and gave off an unpleasant odor. Arrowhead points to the part of the structure that has collapsed.

For molecular analysis, the total genomic DNA was extracted from 24 hr cultures grown on NA using a commercial extraction kit, following the guidelines presented in the HiGene Genomic DNA Prep Kit (Biofact, Daejeon, Korea). The 16S rRNA region was amplified through a polymerase chain reaction (PCR) protocol adapted from Weisburg et al. (1991). To effectively purify the PCR products, the ExoSAP-IT PCR Product Cleaning Reagent (Thermo Fisher Scientific, Waltham, MA, USA) was employed. The sequencing of the 16S rRNA region of the strain KNUB-09-21 was successful, resulting in a fragment of 1,387 base pairs (GenBank no. LC782270). A BLAST search against the NCBI database revealed a striking similarity between the 16S rRNA sequence of KNUB-09-21 and various strains within the genus Pectobacterium, including P. brasiliense, P. carotovorum subsp. carotovorum, and P. versatile, ranging from 99.78% to 99.93%. This comparison definitively established the affiliation of the strain KNUB-09-21 with the genus Pectobacterium. This genetic closeness provides valuable insights into the taxonomic classification of the strain within this specific microbial group.
Recently, a comprehensive analysis was conducted to clarify the classification of Pectobacterium species. This analysis utilized three housekeeping genes (dnaX, leuS, and recA) to refine the taxonomic categorization of various members within the genus Pectobacterium (Portier et al., 2019). In our study, the PCR protocols for amplifying the dnaX, leuS, and recA genes were adapted from Sławiak et al. (2009), Portier et al. (2019), and Waleron et al. (2002), respectively. Using the primer sets outlined in Table 1, the genetic sequences of the strain underwent amplification, subsequently being submitted to GenBank under the corresponding accession numbers LC782271, LC782272, and LC782273. For the alignment of multiple sequences, the software MEGA7 was employed, following the approach outlined by Kumar et al. (2016). The resulting sequences (dnaX, 511 bp; leuS, 486 bp; recA, 734 bp) were utilized for the subsequent phylogenetic analysis with other Pectobacterium species (Table 2).
Table┬Ā1.
Compilation of PCR primers utilized in this study
| Gene | Primer sequence 5ŌĆ▓ŌåÆ3ŌĆ▓ | Reference |
|---|---|---|
| 16S rRNA | Forward: GAG TTT GAT CCT GGC TCA G | Weisburg et al. (1991) |
| Reverse: ACG GCT ACC TTG TTA CGA CTT | ||
| dnaX | Forward: TAT CAG GTY CTT GCC CGT AAG TGC | Sławiak et al. (2009) |
| Reverse: TCG ACA TCC ARC GCY TGA GAT G | ||
| leuS | Forward: TYT CCA TGC TGC CYT AYC CT | Portier et al. (2019) |
| Reverse: TCC AGT TRC GCT GCA TGG TT | ||
| recA | Forward: GGT AAA GGG TCT ATC ATG CG | Waleron et al. (2002) |
| Reverse: CCT TCA CCA TAC ATA ATT TGG |
Table┬Ā2.
GenBank and culture accession numbers of the isolates incorporated in this study
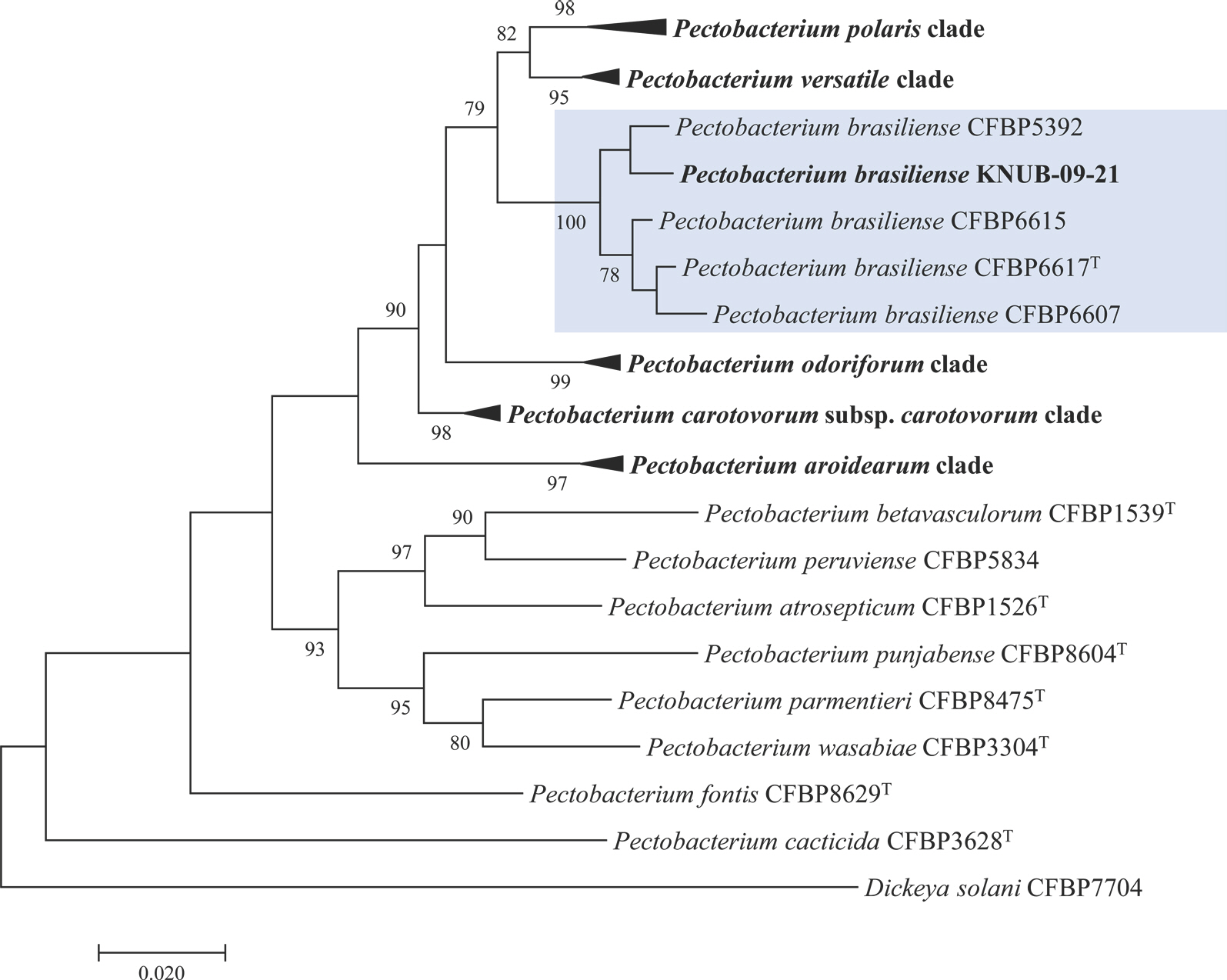
In the construction of the phylogenetic tree, the maximum-likelihood analysis was conducted, employing Kimura's two-parameter model. This was complemented by the application of the nearest-neighbor interchange heuristic search approach, in accordance with the methodology proposed by Felsenstein (1981). The phylogenetic analysis revealed a monophyletic clade uniting the KNUB-09-21 isolate with a set of P. brasiliense strains (CFBP 5392, CFBP 6607, CFBP 6615, and CFBP 6617 T), characterized by a high bootstrap value. This compelling evidence strongly supported their membership within the same species (Fig. 2), thus solidifying the taxonomic relationship and shedding light on the phylogenetic context of the analyzed strains.
Fig.┬Ā2.
Maximum-likelihood phylogenetic tree, constructed from concatenated sequences (dnaX+ leuS+ recA), illustrating the phylogenetic position of Pectobacterium brasiliense KNUB-09-21 among closely related Pectobacterium species. The isolated strain is highlighted in bold. Bootstrap values (based on 1,000 replications) of >70% are displayed at the branch points. Dickeya solani CFBP 7704 was used as the out-group. The scale bar represents 0.020 substitutions per nucleotide position.
The strain KNUB-09-21 underwent an evaluation of compound utilization using the API ID 32 GN system (Biom├®rieux, Marcy l'Etoile, France), following the manufacturer's provided guidelines. The results of this assessment revealed that KNUB-09-21 exhibited positive reactions to several compounds, including N-acetyl-glucosamine, D-glucose, inositol, D-mannitol, D-melibiose, L-rhamnose, salicin, L-serine, and sucrose. Conversely, the isolate demonstrated negative responses to L-alanine, L-fucose, L-histidine, lactic acid, D-maltose, propionic acid, and D-sorbitol. The comprehensive findings from these conventional biochemical assays were in alignment with the results of the molecular analysis, thus affirming the accurate identification of the strain KNUB-09-21 as P. brasiliense (Portier et al., 2019). This harmonization between traditional biochemical assessments and advanced molecular characterization provides robust validation for the strain's taxonomic classification within the context of P. brasiliense.
To fulfill Koch's postulates, pathogenicity assessments were carried out using P. brasiliense KNUB-09-21 on M. charantia plants. The fruits of M. charantia were sterilized with 70% ethanol and subsequently rinsed with distilled water before the inoculation process. Inoculation involved applying a 20 ╬╝l suspension (1 ├Ś 108 cells/ml) of P. brasiliense KNUB-09-21 to M. charantia via syringe injection. Simultaneously, a control group was established by inoculating the plants with a 20 ╬╝l volume of distilled water. The inoculated plants were maintained in a controlled greenhouse environment (28┬░C, relative humidity 80%). After two days, the infected fruits emitted a foul odor, displayed a transition to a yellow color on their surface, and ultimately underwent structural collapse (Fig. 3A). Furthermore, the internal region of the fruit turned dark brown (Fig. 3B). These symptoms observed closely resembled those found in naturally affected M. charantia plants from Goesan. In contrast, the control group did not exhibit any noticeable symptoms (Fig. 3C). Reidentification of the bacterial strain isolated from each symptomatic fruit confirmed its classification as P. brasiliense (data not shown).
Fig.┬Ā3.
Results of the pathogenicity test for Pectobacterium brasiliense KNUB-09-21. (A) The surface exhibited yellowing and collapse. (B) The fruit's interior has darkened to a brown color. (C, D) Momordica charantia inoculated with sterile water (control).

M. charantia has held the distinction of being the largest cultivated crop among subtropical crops in Korea, surpassing various other fruits and vegetables (Jeong et al., 2020). This crop is susceptible to a range of bacterial, fungal, and viral pathogens, including Colletotrichum gloeosporioides and Rhizopus stolenifer (Kim et al., 2015; Kwon and Jee, 2005). Notably, while bacterial soft rot diseases have been reported globally, there have been no documented cases of bacterial diseases affecting M. charantia in Korea. Bacterial soft rot, induced by pectinolytic soft rot bacteria belonging to the genera Pectobacterium and Dickeya, is considered one of the most destructive diseases of vegetables worldwide (Ma et al., 2007). The genus Pectobacterium, known for its wide host compatibility, can trigger soft rot symptoms in various plant species due to its ability to produce enzymes that degrade plant cell walls (Hugouvieux-Cotte-Pattat et al., 2014; Maung et al., 2022). The use of phylogenetic analysis involving housekeeping genes (dnaX, leuS, and recA) and investigations into compound utilization has proven effective in classifying Pectobacterium species (Portier et al., 2019).
Our study, utilizing a comprehensive investigative approach that includes analyses of the 16S rRNA region sequence, multilocus sequence, and thorough evaluation of biochemical traits, definitively identifies P. brasiliense as the causative agent of the observed soft rot in M. charantia in Korea. Despite recent advancements in pathogen diagnostic methods for soft diseases that allow for sensitive and specific detection of target pathogens (Jin et al., 2022), there is a lack of comprehensive investigation regarding the survey of bacterial pathogens in M. charantia. This underscores the need for further investigation of potential bacterial diseases that can affect M. charantia. Our findings provide valuable insights that can contribute to the development of effective plant pathology strategies. Importantly, they lay a solid foundation for designing targeted control measures to combat soft rot and mitigate associated economic losses caused by this identified phytopathogen. In conclusion, our results provide foundational data that can inform future plant pathology efforts, offering a promising direction for effective disease management.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






