



Introduction
Lily is one of the main flowers in South Korea, influencing the revenue importantly as the most exported flower (Song and Kim, 2021). Most lily flowers produced in South Korea were exported to Japan where 90% of imported lilies were from Korea (Choi et al., 2018; Kim and Kim, 2015). A total of 398 tons of lily flowers produced in Korea were exported to Japan in 2021, with an annual income reaching about 4 million dollars (Korea Agro-Fisheries and Food Trade Corporation, 2021).
Lily bulbs cultivated in Korea were mostly imported from the Netherlands (Jang and Kim, 2016). Recently, lily fasciation caused by Rhodococcus fascians has been reported in some lily grown in greenhouses in South Korea. Constrainedly, 1.3 million lily bulbs from the Netherlands were abrogated because of infection by R. fascians, a quarantine bacterium, in 2020 (Lim et al., 2021). In Jeju Island, one of the major regions cultivating lily in Korea, strict quarantine is absolutely necessary to prevent bacterial fasciation caused by R. fascians in lily.
The actinobacterium R. fascians is a gram-positive bacterium that has a broad range of hosts. It infects not only dicotyledonous plants, but also some monocotyledon plants such as Liliaceae (Francis et al., 2016). Symptoms of infected plants by R. fascians include leafy gall formation, leaf deformation, multiple shoot formation, loss of apical dominance, and abnormal development caused by unbalanced phytohormones (Goethals et al., 2001; Nikolaeva et al., 2009; Putnam and Miller, 2007; Stes et al., 2013). To detect R. fascians, virulent plasmid genes such as att and fas could be targeted generally for polymerase chain reaction (PCR) (Savory et al., 2017; Serdani et al., 2013). Virulence gene vicA in chromosome can also be used for diagnosis of R. fascians. However, all Actinobacteria could be amplified with vicA marker (Nikolaeva et al., 2012; Savory et al., 2017). Analyzing 16S rRNA sequences would not be able to detect R. fascians at species level (Dhaouadi and Rhouma, 2020; Patel et al., 2004).
In this study, lilies grown in greenhouses located in Jeju Island were collected from 2021 to 2022 to detect target bacteria R. fascians. Two different sets of PCR primers were used to diagnose R. fascians from bacteria isolated from lily bulbs. One set of primers targeted virulent fasA gene in the plasmid which is essential for symptom development (Savory et al., 2017). And the other set of primers targeted a putative glycosyltransferase epsH gene in chromosome. To investigate whether bacterial isolates could infect lily leaves directly or not, pathogenicity assay was carried out using lily leaves and bacteria identified as R. fascians. Furthermore, bacteria were re-isolated from inoculated tissues of wounded lily leaves.
Materials and Methods
Isolation of R. fascians
Lilies with symptoms similar to those infected by R. fascians were collected from five greenhouses located in Jeju Island during harvest season in 2021 and 2022. Lily cultivars were Jambesi, Jojia, Swera, Rubina, Siberia, and Wooritower. Symptomatic lilies were collected and placed in plastic bags with 100% humidity until bacterial isolation. The bacteria were isolated according to the method by Park et al. (2022). The lily bulb, stem, and leaf tissues were cut in to a size of 0.5 cmx0.5 cm using a razor. Cut samples were sterilized with 1% sodium hydrochloride and then 70% ethanol for 30 sec each and finally rinsed with sterile water three times for 1 min each. In order to isolate bacterial cells from plant tissues, sterilized tissues were vortexed in a microtube filled with 1.0 ml sterile water for 1 hr. To isolate bacterial cells, 100 µl of the plant tissue maceration was spread onto a tryptic soy agar media (TSA; Becton, Dickinson and Company, Claix, France), using a sterilized loop, and incubated at 28 o C for 2 days until bacterial colonies appeared. Smooth and yellow colonies (typical growth shape and color of R. fascians) were selected and further incubated for molecular diagnosis.
Identification of R. fascians by molecular diagnosis.
To harvest mass of bacteria, bacterial isolates were cultured in tryptic soy broth (Becton, Dickinson and Company) media at 28 o C in a shaking incubator at 3,000 × g for 24 hr. The bacterial suspension was centrifuged at 3,000 ×g for 3 min. Bacterial pellet was then used for DNA extraction with a DNeasy blood & tissue kit (Qiagen, Hilden, Germany) using gram-positive bacteria extraction protocol. Total DNA of the bacterial isolate was amplified with two sets of specific primers. One set of primers targeted P450 of the pathogenic plasmid and the other set of primers targeted RS02785 of chromosomal DNA (Table 1). PCR cycling conditions for P450 were as follows: 94 o C for 5 min; 30 cycles of 94 o C for 30 sec, 55 o C for 30 sec, and 72 o C for 1 min; and 72 o C for 5 min. For PCR with the other set of primers targeting RS02785, the annealing temperature was 60 o C. Other PCR conditions were the same as those used for P450. Amplified DNA was confirmed by 1% agarose gel electrophoresis at 70V for 60 min. PCR products were then sequenced by Macrogen (Seoul, Korea). Sequences were then analyzed using BLAST service of NCBI.
Table 1
Primer sequences used for identification of Rhodococcus fascians and target locus of R. fascians amplified by each primer set
Pathogenicity assay of the bacterial isolates
Plant materials: Lily bulbs (Lilium longiflorum cultivar Siberia) were planted in plastic pots (Ø10 cm) containing commercial soil (Number-One, Hongseong, Korea) and 10 % perlite (Parat, Sam Son, Seoul, Korea) and incubated at 28 o C with light intensity at 8,000 lux for 14 hr of day time for four weeks until fourth leaves grew completely.
Bacterial inoculum: Bacterial strain isolated from the greenhouse in Jeju Island and identified as R. fascians 3AF-1 was used in the in vivo test. In order to make bacterial inoculum, the bacterial isolate was cultured on TSA media and incubated at 28 o C for 2 days. To harvest bacterial cells, 10 ml of sterilized water was added onto the bacteria grown plate and rubbed with a loop. The concentration of bacterial suspension was adjusted to 1.0×108 cfu/ml using a spectrophotometer (Optizen POP, Cheongju, Korea). It was then used as an inoculum.
Inoculation and re-isolation of R. fascians: Completely grown lily leaves were used for inoculation in this bio-test. Four sites of leaves were scarred with sterilized needles and inoculated on the wounded sites with the bacterial inoculum. Unwounded lily leaves were also inoculated. Both inoculated leaves were incubated in a dew chamber (DA-DC, DONG-A, Siheung, Korea) at 28°C with relative humidity of 100%. After 1 day, plants were moved into a plant chamber at 28°C with 90% relative humidity and 14 hr of light time for 4 days until the symptom appeared.
To confirm the bacterial isolate as R. fascians, bacterial isolates were re-isolated from inoculated leaves. Leaves were cut at 0.5 cm×0.5 cm and sterilized the same as the first isolation. Re-isolated bacterial cells were incubated on TSA at 28°C until bacterial colony appeared. Total DNA was extracted from re-isolated bacterial cells and amplified with specific primer RS02785 for identification.
Results and Discussion
Bacterial isolation
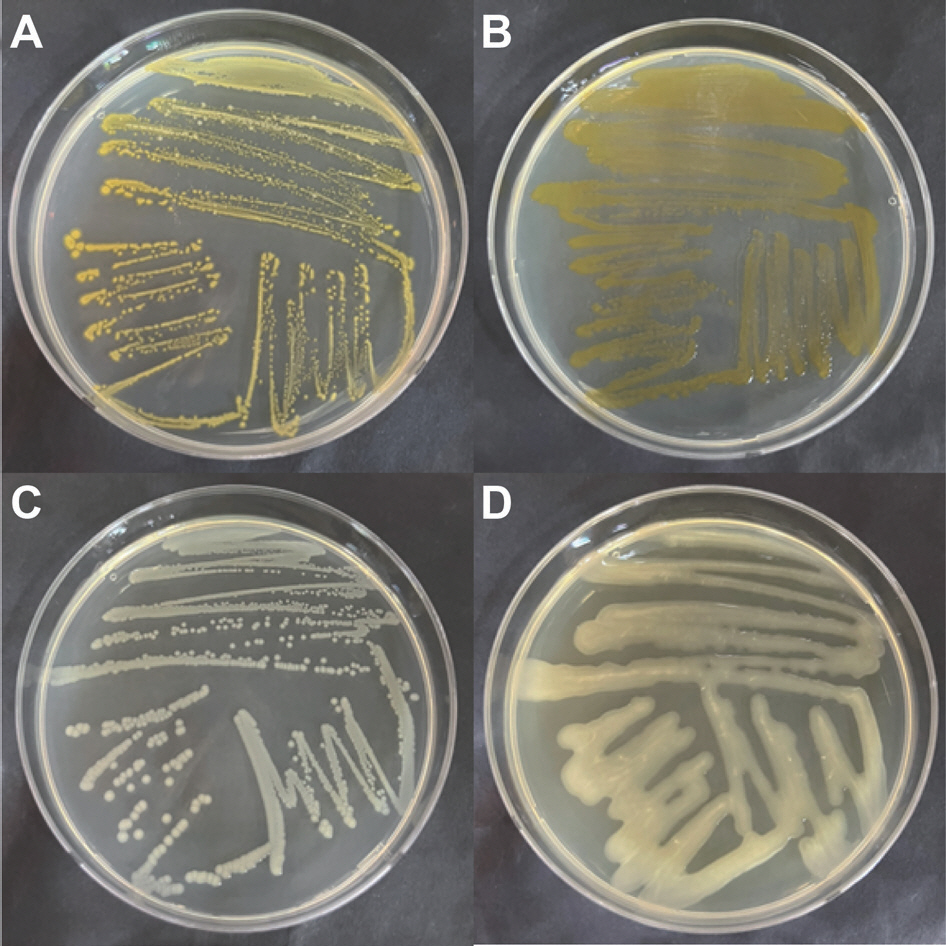
To diagnose causal bacteria from infected lilies showing symptoms suspected as lily fasciation, a total of 508 bacteria were isolated from lily greenhouses in this study. Among them, 176 bacterial colonies showing either yellow or smooth type were selected for DNA test (Fig. 1). Colony types of R. fascians on TSA have already been reported. Colony of R. fascians containing carotenoid pigment showed orange color (Stes et al., 2011). In the initial stage, colonial shape and color of the bacteria have been reported which was smooth in shape and creamy in color that would turn to yellow (Dhaouadi and Rhouma, 2020).
Molecular analysis of bacterial isolates
Two different primer sets for diagnosing bacterial isolates were compared in this study in order to find a more applicable primer set in practice. Using primer P450 fasA gene in the plasmid was amplified in size of 538 bp whereas a putative glycosyltransferase epsH gene was amplified 258 bp of chromosome. As a result of PCR analysis, three bacterial DNA samples were amplified using the set of P450 primers targeting bacterial plasmid (Fig. 2). A total of 12 bacterial isolates were amplified using RS02785 primer sets, including the three bacterial isolates identified as R. fascians with P450 primer set (Fig. 3). Based on blast data, all 12 bacteria amplified with a primer set of RS02785 were identified as R. fascians.
Fig. 2.
Polymerase chain reaction product amplified with specific primer P450 targeting FasA gene in plasmid o f Rhodococcus fascians. Lanes: 1-kb DNA ladder (BIOFACT, Co. Ltd.) (M), R. fascians (Rf), bacterial strains isolated from lily bulbs (18, 22, and 23). The arrow indicates 538 bp.
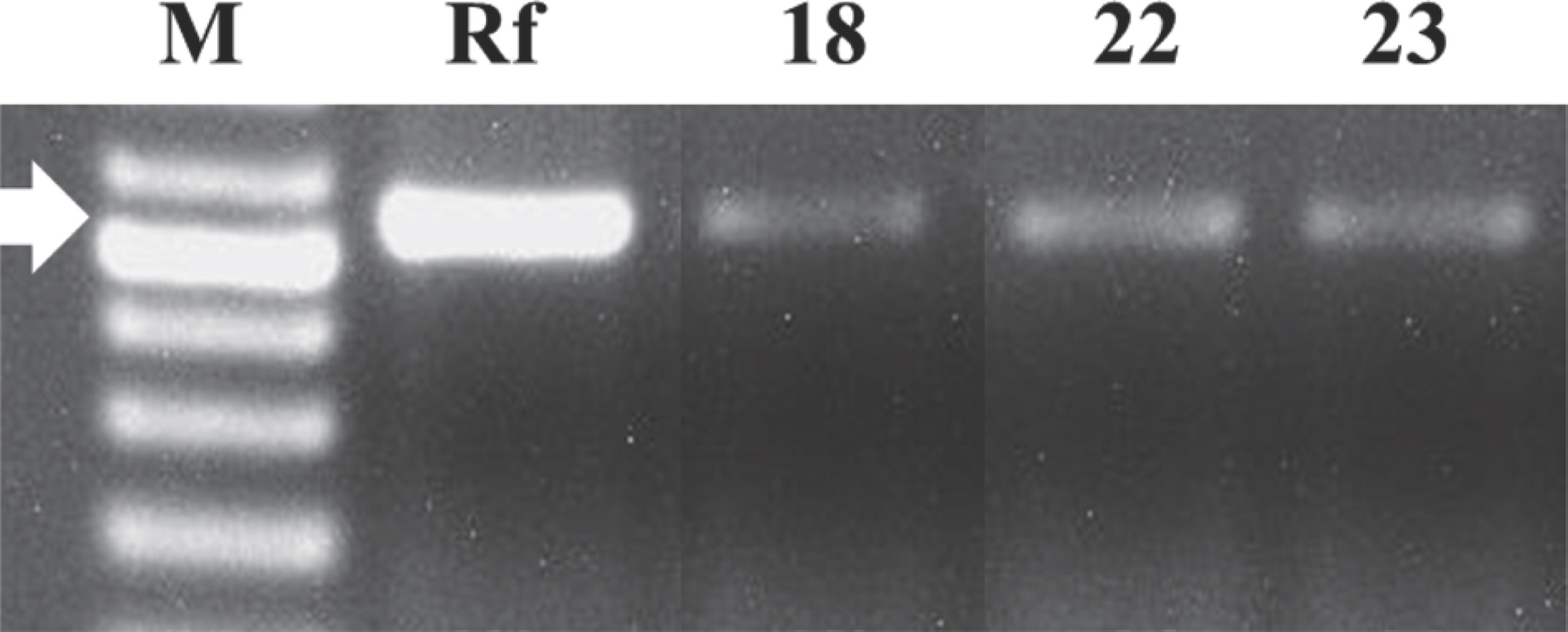
Fig. 3.
Polymerase chain reaction product amplified with specific primer RS02785 targeting chromosome of Rhodococcus fascians. Lanes: 1-kb DNA ladder (BIOFACT, Co. Ltd.) (M), R. fascians (Rf), bacterial strains isolated from lily bulbs (18--156). The arrow indicates 258 bp.

Using P450 primers for diagnosis of R. fascians has been reported by targeting the FasA gene in plasmid carrying pathogenicity (Pertry et al., 2010; Serdani et al., 2013). Plasmid pFID188 is related to recovery of the pathogenicity of R. fascians. The sequence of fasA gene has 100% identity with the partial gene sequences of pFID188 (Crespi et al., 1992; Stamler et al., 2015). In our study, nine bacterial isolates not amplified with P450 primers would not include pathogenic fasA gene. However, they could be reintroduced with the pFID188 plasmid by other pathogenic wild type pathogenic bacteria, resulting in recovery of its pathogenicity. Therefore, using both primer sets may be recommended for complete quarantine of this bacterial disease.
Bio-test for pathogenesis of bacterial isolates
Inoculation onto lily leaves: To be certain that isolated bacteria were causal bacteria from the first infected lilies, isolated bacteria identified as R. fascians using RS02785 were inoculated onto lily leaves. As a result of bacterial inoculation, no symptom was observed on lily leaves without wound (data not shown). However, apparent symptoms were observed as black gray on wounded leaves inoculated with the bacterial suspension (Fig. 4). No symptom appeared on wounded or unwounded leaves (treated with H2 O instead of bacterial suspension).
Fig. 4.
Lily plants treated with H2 O (A) and inoculated with Rhodococcus fascians 3AF-1 (B) after wounding. Plants were taken at 4 days after bacterial inoculation. Concentration of the bacterial suspension was 1.0×108 cfu. Square boxes are magnified symptoms (red circles).

In this result, bacteria could not infect plant leaves directly, suggesting that these bacteria might be non-pathogenic ones. Although bacterial strains were isolated from lily leaves showing fasciation, it seemed bacterial isolates were not virulent enough to infect lily leaves. One study has shown that a less virulent R. fascians strain could form leaf gall symptom on Nicotiana tabacum cv. Havana and N. tabacum cv. Xanthi leaves (Cabecinhas, 2014). For pathogenesis of R. fascians, FasA gene in bacterial plasmid is required for efficient infection of host plant (Crespi et al., 1994; Pertry et al., 2010). The pathogenic plasmid including FasA or FasD could be introduced to non-pathogenic bacteria which could acquire pathogenicity and infect the host plants (Lawson et al., 1982; Stange et al., 1996).
Re-isolation of bacterial isolates: Lily leaves inoculated with a bacterial strain suspected as R. fascians were sampled for bacteria isolation to confirm that these bacteria were the same as the inoculum. Bacterial colonies were not observed on untreated leaves. However, abundant bacterial colonies were isolated from inoculated lily leaves (Fig. 5). DNA of the isolated bacteria were amplified using RS02785 primer sets and identified as R. fascians (Table 2). Some studies have shown that non-virulent bacterial strains could infect plants by wounding. R. fascians deficient in its pathogenic plasmid could not directly infect plant cells, but can proliferate on epidermal leaves (Cornelis et al., 2001). On tobacco plants, symptoms of fasciation caused by R. fascians have been observed on wounded leaves (Cabecinhas, 2014). Furthermore, avirulent bacteria could affect plants as virulent bacteria. However, colonization by the avirulent bacteria was lower than that by virulent ones (Dhandapani et al., 2018). Similarly, many R. fascians can grow at wound sites of tobacco plants (Cornelis, 2000). In our previous study, numbers of bacterial colonies were isolated on TSA media from inoculated leaves with wounding. These results indicate that R. fascians can infect through wounding site and proliferate in leaves. Therefore, R. fascians isolated in Jeju Island could not infect lily plant directly, but it could infect lily through wound by insects or artificial pruning.
Fig. 5.
Numbers of colonies on tryptic soy agar medium incubated with the suspension extracted from lily tissues treated with H2 O and inoculated with Rhodococcus fascians 3AF-1 after wounding.
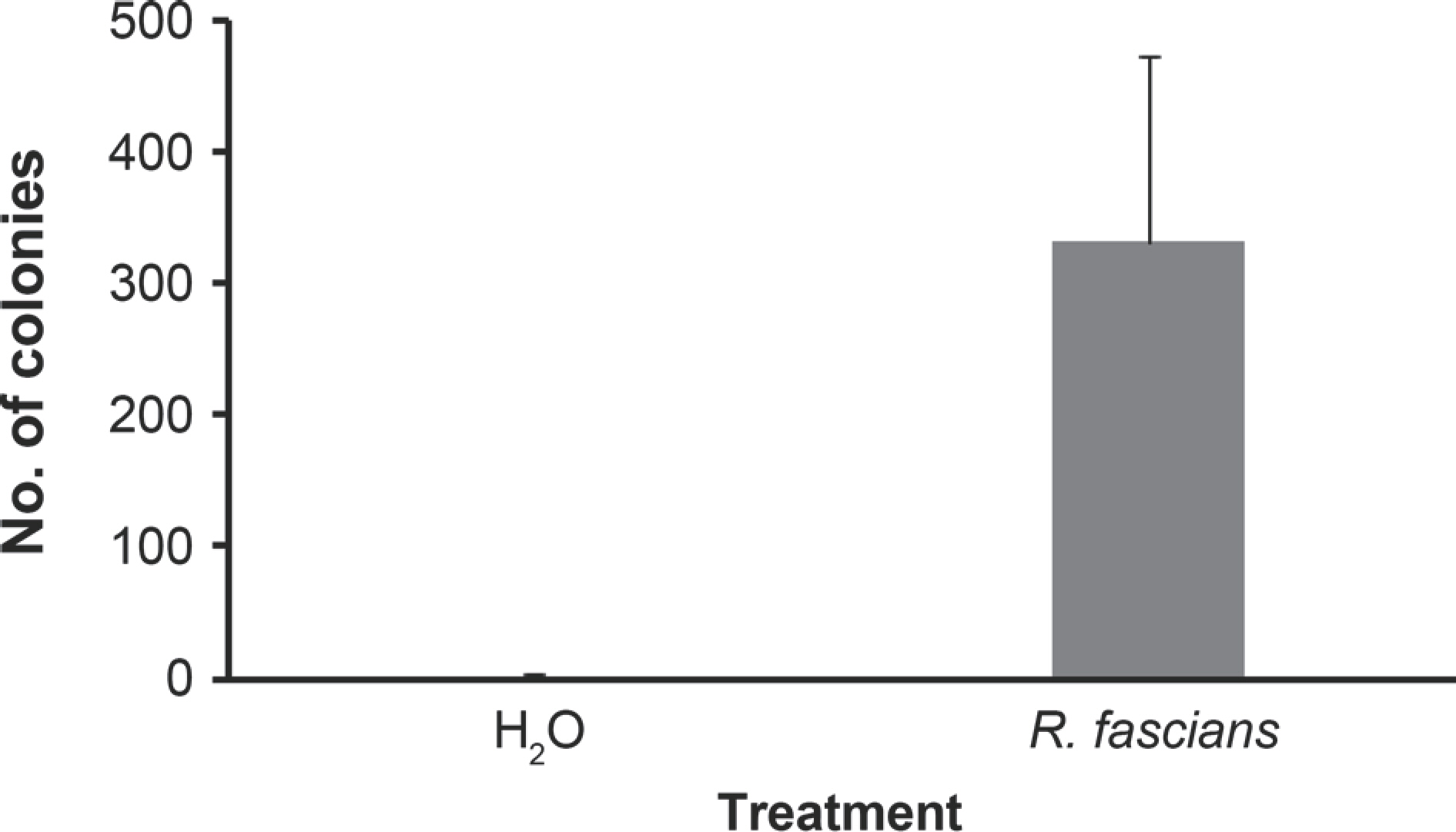
Table 2
Sequences analysis of PCR products of bacterial strains re-isolated from lily leaves inoculated with an artificial wound inoculation solution a
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






