



서 론
식용 버섯은 칼로리와 지방의 함량이 낮고 단백질, 식이섬유, 미네랄 함유가 높아 영양적 측면뿐만 아니라 항산화 효과로 건강식품으로 널리 활용되고 있다(Bernaś 등, 2006; Lu 등, 2020; Rangel-Vargas 등, 2021). 이러한 식용 버섯 중 목이버섯(Auricularia auricula-judae)은 아시아에서는 한약재로 사용되어왔으며, 독특한 식감과 향으로 우리나라를 비롯하여 일본, 중국, 대만 등에서 식품으로 식용되고 있다. 목이버섯의 분류체계는 담자균류(Basidiomycota), 목이목(Auriculariales), 목이과(Auriculariaceae), 목이속(Auricularia)에 속하며, 전 세계적으로 분포한다. 특히 중국, 일본, 우리나라에서 양적으로 많은 생산과 소비가 이루어지고 있다(Chen과 Hou, 1978).
목이버섯의 재배는 통나무나 멸균된 톱밥을 이용하는 표고버섯(Lentinula edodes)의 재배 방법과 유사하다(Tu와 Wu, 1989). 상업적으로 재배되는 버섯 생산에 사용되는 배지는 버섯이 판매될 때까지 충분한 영양분을 제공하지만 이 영양분은 부생성 곰팡이에게도 선호적인 영양분을 제공하기 때문에 버섯 종균과의 경쟁으로 인해 재배에 어려움이 있다(Houbraken 등, 2019; Kim 등, 2019; Kosanović 등, 2015; Lee 등, 2017; O'Brien 등, 2014; O'Brien 등, 2017; Radványi 등, 2016). 특히 버섯의 3차 구조 형성이 시작되면 공기와 차단하여 기르던 2차균사를 공기 중으로 노출시키게 되는데 이 기간은 부생성 곰팡이가 배지에 감염하여 포자를 형성하는 기간보다 훨씬 길어 버섯은 배지의 영양분을 이용하지 못하게 된다.
버섯 산업을 방해하는 이러한 경쟁적인 부생 곰팡이로는 Trichoderma spp., Penicillium spp., Aspergillus spp. 등이 보고되었다(Šašić Zorić 등, 2023). 이 중 Trichoderma spp.는 푸른곰팡이(green mold)로도 알려져 있는데 버섯 배지에 감염을 일으키는 주요 곰팡이로 재배업자에게 경제적 피해를 준다(Kim 등, 2019; Šašić Zorić 등, 2023). Kim 등(2019)은 표고버섯 톱밥 배지에 피해를 주는 곰팡이를 조사하였는데, 총 405개의 부생 곰팡이를 분리 및 동정한 결과 Penicillium spp. (26.2%)와 Trichoderma spp. (9.5%)가 우점한다고 보고하였다. 특히 Trichoderma harzianum의 경우 수집한 모든 농장에서 분리되어 가장 흔히 발견되는 배지 감염균으로 보고하였다(Kim 등, 2019).
유럽과 북미의 버섯 산업에 경제적으로 가장 중요한 버섯인 양송이 버섯(Agaricus bisporus) 재배에서도 부생 곰팡이는 큰 손실을 불러온다. 대표적으로 Trichoderma aggressivum이 보고되었는데, 이 종은 양송이 버섯 재배에서 치명적인 수량 손실을 초래하기도 하였다(Kosanović 등, 2015; O'Brien 등, 2014; O'Brien 등, 2017; Radványi 등, 2016). 이와 다르게 Houbraken 등(2019)은 양송이 재배에 가장 큰 문제 감염균으로 Penicillium hermansii를 보고하였으며, 미국, 네델란드, 영국의 가장 큰 경제적 피해를 주는 푸른곰팡이 종으로 규명하였다.
국내 배지 이용 버섯 재배 선호도 증가에 따른 버섯 산업의 지속적 성장에도 불구하고 경쟁 곰팡이 감염균은 여전히 버섯 산업에 가장 위협적인 문제이다. 특히 버섯은 친환경 재배가 선호적이며, 2019년부터 시행된 농약허용물질목록관리제도(Positive List System, PLS)에 따라 잔류허용 기준에 대한 경각심으로 농약 사용이 제한적이다.
생물학적인 방제제(biological control agents, BCA)는 일반적으로 공동배양법(dual-culture method)을 이용하여 방제하고자 하는 병원체에 대하여 생물학적 방제제를 함께 공동배양하여 길항성의 여부를 정량적으로 탐색한다. BCA로 사용되는 대표적인 세균들은 Pseudomonas spp., Bacillus spp.를 비롯하여 Streptomyces spp. 등 다양하다(Bonaterra 등, 2022; Boukaew 등, 2011; Chamberlain과 Crawford, 1999; Trejo-Estrada 등, 1998). 이 중 Streptomyces spp.는 다른 세균들과 다르게 기중균사가 발달한 뒤 포자가 형성되기 시작할 때 다양한 2차 대사산물이 생산되는 특징이 있어, in vitro 실험을 수행할 때 빠르게 생장하는 곰팡이종의 경우 길항 효과를 명확히 구분하기 어려울 수 있다(Bonaterra 등, 2022).
이 부분을 보완할 수 있는 방법 중 하나는 Kunova 등(2016)에 의해 제안된 방법이다. 이 방법은 대상 곰팡이의 성장 속도를 고려하여 빠르게 자라는 병원성 곰팡이의 경우 Streptomyces spp.의 길항성을 검정하기 위해 배지에 3일 전에 접종하여 기중균사와 포자를 형성하게 하고, 그 이후 병원성 곰팡이를 접종하는 공동배양법으로 효율적으로 길항세균을 선발할 수 있다고 보고하였다(Kunova 등, 2016).
이 연구의 목적은 목이버섯 배지에 부생균에 의해 감염된 피해 농가로부터 1) 원인이 되는 감염 곰팡이를 분리하고, 2) 분리한 감염 곰팡이균의 형태 및 분자생물학적 동정을 통해 우점균을 밝히고, 마지막으로 3) 친환경적인 방제제로서의 Streptomyces 종의 적용을 통해 잠재적인 친환경 방제제를 선발하는 것을 목적으로 하였다.
재료 및 방법
경쟁 곰팡이 오염 목이버섯 배지 수집 및 병원균 분리.
2021년 10월 장흥 버섯농가(34°65’86.04” N, 126°95’89.58” E)와 2021년 11월 전남 순천시 버섯농가(34°84’43.30” N, 127°45’57.37” E)에서 경쟁 곰팡이에 오염된 배지를 4개 및 10개를 각각 수집하였다.
수집된 총 14개의 버섯 배지의 감염된 세 개 구획을 랜덤하게 선택하였다. 선택된 배지에 멸균된 루프를 이용하여 오염된 경쟁 곰팡이균을 채취하여 1,000 µg/ml 스트렙토마이신(strep-tomycin)이 첨가된 물한천배지(20 g agar, 1 l)에 접종하여 25°C 암상태의 항온기에서 3일 동안 배양하였다. 자라나온 경쟁 곰팡이 균사는 5×5 mm로 균사 말단을 잘라 새로운 감자한천배지(potato dextrose agar [PDA] media; Difco Laboratories, Detroit, MI, USA)로 옮겨 원인 곰팡이균을 확보하였다. 확보된 배양 균주는 단포자 분리를 통해 순화한 뒤, 장기 보존을 위해 포자를 20% Glycerol 용액에 희석하여 -80°C 초저온냉동고에 보관하였다.
균주의 형태학적인 특성.
분리 균주의 형태학적인 특성을 확인하기 위해 9 cm, PDA 배지에서 7일 배양하였다. 배지의 앞과 뒷면에 나타난 균총의 모양, 색깔 등의 형태적 특징을 육안으로 살펴보았다. 포자경과 포자는 Zeiss Axio Imager A1 현미경(Carl Zeiss, Oberkochen, Germany)으로 관찰하였다.
Genomic DNA 추출 및 염기서열 분석.
분자생물학적 동정을 위해 감염 곰팡이균으로부터 genomic DNA를 추출하였다. PDA 배지에서 성장한 균사체를 5×5 mm로 자른 뒤, 3-5 mm 의 bead(유리구슬)가 5-10개 첨가된 2 ml tube에 넣었다. 그 후 KCl buffer (100 mM Tris-Cl [pH 8.0], 10 mM EDTA [pH 8.0], 1 M KCl) 300 µl를 넣어 bead beater (BioSpec mini bead beater 8; BioSpec, Bartlesville, OK, USA)를 이용하여 균사를 마쇄하였다. 이후 과정은 이전 연구에 기술한 방법대로 DNA를 추출하였다(Chi 등, 2009). 추출된 DNA는 최종적으로 TE buffer (10 mM Tris-Cl [pH 8.0], 1 mM EDTA [pH8.0]) 100 µl에 용해시킨 뒤, NanoDrop ND-1000 spectrophotometer (NanoDrop Technologies, Inc., Wilmington, NC, USA)를 통해 DNA의 양을 측정하였다. 추출된 DNA는 실험 사용 전까지 -20° C에 보관하였다.
곰팡이 종 동정을 위해 internal transcribed spacer (ITS) 영역을 polymerase chain reaction (PCR) 증폭한 뒤 염기서열 정보를 확보하였다. Primer는 forward primer로 ITS1F(5’-TCCGTAG-GTGAACCTGCGG-3’) (Gardes와 Bruns, 1993)과 reverse primer 로 ITS4(5’-TCCTCCGCTTATTGATATGC-3’)을 사용하였다(White 등, 1990). PCR 반응을 위한 준비는 다음과 같이 수행하였다. 우선 genomic DNA (25 ng) 2 µl, forward primer와 reverse primer를 10 pmol/µl로 희석하여 각각 1 l씩 넣고, 2× PCR Mas-ter mix solution (iNtRON Biotechnology, Sungnam, Korea) 10 µl 그리고 마지막으로 멸균증류수(sterilized distilled water)를 넣어 총 용량 20 µl가 되도록 준비하여 PCR을 수행하였다.
이 실험에서 우점종으로 분리된 Trichoderma 속균의 대표균주 선발을 위해서 RNA polymerase II second largest subunit (RPB2) 영역의 염기서열을 확보하였다. Primer는 forward primer로 fRPB2-5F(5’- GAYGAYMGWGATCAYTTYGG-3’)과 reverse primer로 fRPB2-7CR(5’- CCCATRGCTTGYTTRCCCAT-3’)을 사용하였다(Liu 등, 1999). PCR instrument (Applied Biosystems, Thermo-Fisher Scientific, Marsiling, Singapore) 반응 조건은 최초 94°C에서 3분 denaturation한 뒤, 94°C에서 30초, 52°C에서 30초, 72°C에서 1분 과정을 반복적으로 총 30회 반복하였다. 마지막으로 72°C에서 5분간 증폭하였다.
PCR의 증폭 여부를 확인하기 위해 DNA 염색 시약(EcoDye™ DNA Staining Solution; SolGent, Daejeon, Korea)이 첨가된 0.7% agarose gel을 이용하여 전기영동하였다. 염기서열 분석(Bioneer, Daejeon, Korea) 후 확보된 정보는 National Center for Biotechnology Information (NCBI)의 Basic Local Align-ment search Tool (BLAST) (https://blast.ncbi.nlm.nih.gov/Blast.cgi)을 통해 데이터베이스의 염기서열과 상동성이 가장 높은 종의 정보와 비교하였다.
목이버섯 종균과 경쟁 감염 곰팡이균의 균사생장 비교.
목이버섯 종균과 분리된 감염 곰팡이의 생장 조사를 위해 균사의 가장자리를 직경 5 mm cork borer를 이용하여 확보한 뒤, 9 cm PDA 배지의 중앙에 접종하였다. 온도 범위는 15-25°C 범위를 5°C 간격으로 나누어 배양기에서 배양하였다. 균사생장의 차이는 24시간, 48시간, 72시간 배양 후 균사생장을 측정하였다.
감염 경쟁 곰팡이병균에 대한 길항세균 Streptomyces sp. 균주 선발.
길항세균으로 사용할 6종의 Streptomyces sp. 균주들은 국립생물자원관으로부터 분양받아 실험에 사용하였다. 목이버섯 배지에서 분리된 감염 곰팡이인 Trichoderma sp.와 Penicillium sp.와의 공동배양(dual culture) 방법을 통해 수행하였다. 모든 실험은 9 cm, PDA 배지에서 수행하였다.
감염 균주 Trichoderma spp.와 Penicillium spp.는 PDA 배지에 접종한 뒤 25° C에서 4일간 배양하여 실험에 사용하였다. 배양 균주의 균사 말단 부위를 직경 5 mm로 접종원을 준비하였다. 길항세균으로 사용할 Streptomyces spp.는 PDA 배지 전면에 streaking한 뒤 28° C에서 7일간 암상태로 배양하여 동일하게 균사 부위를 직경 5 mm로 준비하여 실험에 사용하였다.
공동배양은 9 cm PDA 배지의 중앙에 경쟁 곰팡이 Trichoderma spp.와 Penicillium spp.의 준비한 직경 5 mm 균사를 중앙에 접종하였다. Streptomyces spp.는 중앙으로부터 3 cm 떨어진 곳에 삼각형 모양으로 3곳에 접종하였다. 음성 대조군으로 6 mm paper disc를 놓은 뒤 멸균증류수 15 µl를, 양성 대조군으로 베노밀 수화제(a.i. 50%, WP)를 1,000배로 희석하여(1 g/l) 15 µl를 처리하였다. 25°C 배양기에서 3일간 배양한 뒤 저지대를 측정하였다. 모든 실험은 3반복으로 수행하였으며, 총 3번의 독립적인 실험을 수행하였다.
심층(in-depth) 분석을 통한 효과적인 Streptomyces sp. 균주 선발.
일반적으로 사용되는 공동배양법을 통해 선발된 Streptomyces spp. 균주들의 심층 분석 실험을 수행하였다. 심층 분석은 Kunova 등(2016)에 의해 수행된 공동배양법을 수정하여 적용하였다. 간략하게 설명하면 다음과 같다. 접종원으로 사용할 길항세균 Streptomyces sp. 균주들을 28° C, 7일간 배양하여 공동배양할 9 cm PDA 배지의 중앙에 cell scraper (90020; SPL TM, SPL Life Sciences Inc., Pocheon, Korea)를 이용하여 1 cm 두께로 접종하였다. 이후 28°C, 3일간 배양하였다. 3일 뒤, 4일간 배양된 감염균(Trichoderma sp.과 Penicillium sp.)의 말단 부위를 직경 5 mm로 준비하여 미리 접종된 Streptomyces 종들이 자라고 있는 PDA 배지의 양쪽 끝에 접종하였다. 대조구는 멸균된 여과지(80×7 mm)를 9 cm PDA 배지의 중앙에 놓은 뒤, 멸균증류수와 베노밀 수화제(1 g/1 l)를 각각 200 µl를 떨어뜨린 뒤, 감염균을 바로 접종하였다. 감염균 접종 4일 후 균사 저지대를 측정하였다. 모든 실험은 3반복으로 수행하였으며, 총 3번의 독립적인 실험을 수행하였다.
결과 및 고찰
목이버섯 푸른곰팡이병균의 분리 및 동정.
장흥 버섯농가에서 감염배지 4개로부터 10개의 균주를 분리하였다. 순천 버섯농가에서 감염배지 15개로부터 44개를 분리하여 총 54개의 균주를 확보하였다(Fig. 1, Table 1). 균학적 특성을 검정하기 위해, PDA 배지에서 7일간 배양 후 배지의 앞면과 뒷면의 균사 생장, 색, 형태 등을 관찰하였다. 분리된 감염 균주들의 대부분은 PDA 배지에서 하루가 지나면 백색의 균사가 방사선상으로 빠르게 자라며 시일이 지남에 따라 녹색에서 짙은 녹색을 띄었다. 배지의 뒷면은 짙은 녹색과 회색에 이르는 색을 관찰할 수 있었다(Fig. 2A, B).
Fig. 1.
Symptoms of naturally-contaminated wood ear mushroom (Auricularia auricular-judae) farm infected with blue-green mold and contaminated media. (A) Wood ear mushrooms farm infected with blue-green mold. (B) Discarded sawdust media. (C, D) Close-up view of media contaminated with blue-green mold. (E) Early (left two panels) and late (right two panels) contamination of sawdust media with blue-green mold. The upper side of the pictures in (E) shows the infected appearance on the top side of the sawdust media, and the bottom provides a side view.

Table 1.
List of collected isolates, locality, species and GenBank accession numbers for ITS region sequences
Fig. 2.
Morphological characteristics of three representative Trichoderma sp. and three Penicillium sp. (A, B) The front (top) and reverse sides (bottom) of 7-day-old colonies on potato dextrose agar (PDA) medium. (C, D) Conidiophores after 2 weeks of growth on PDA. (E, F) Conidia. Scale bars: C-F=10 μm.
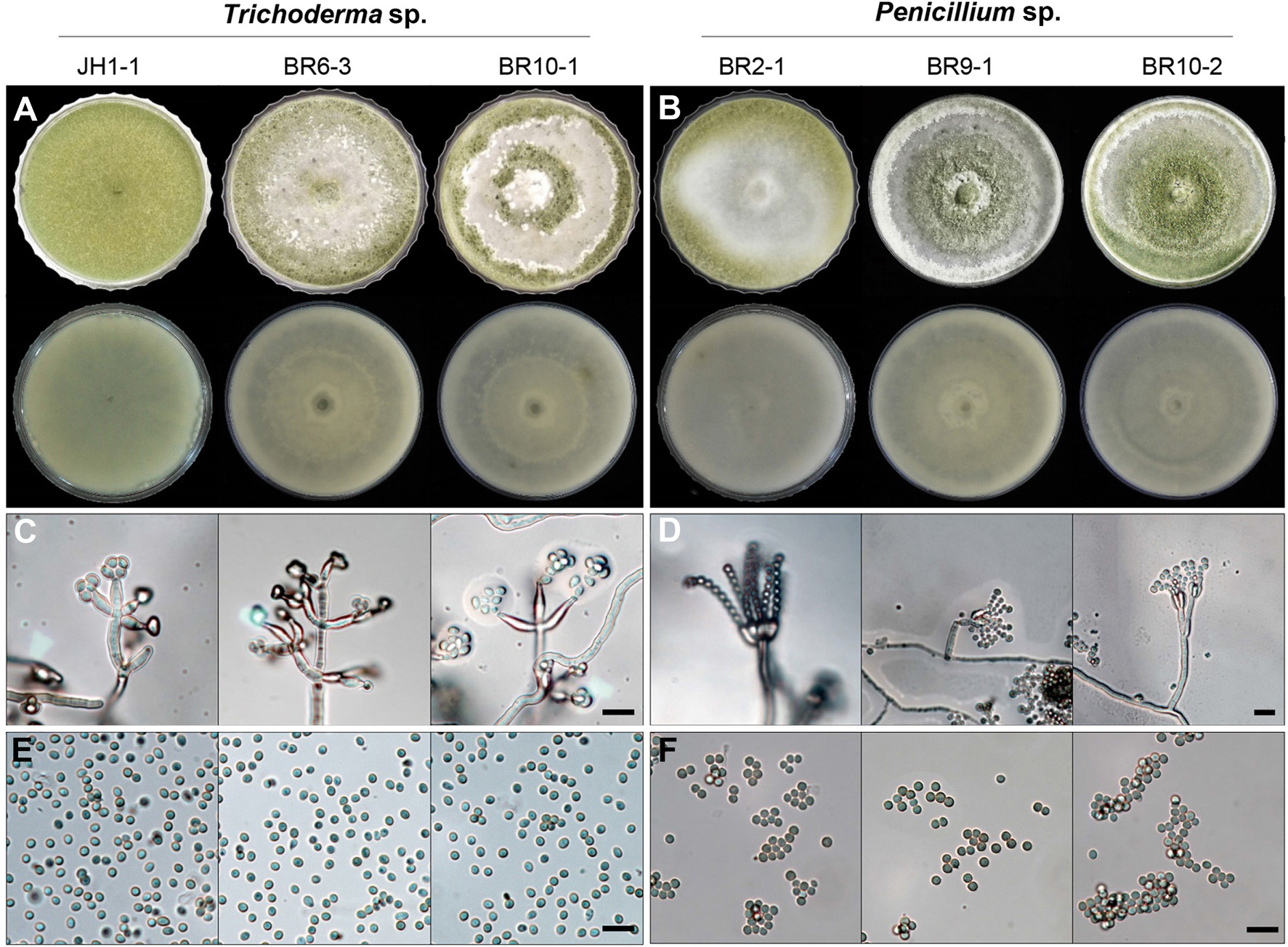
현미경을 통한 관찰 결과 분리된 모든 균은 균사의 격막이 관찰되었으며, 분생자경(Fig. 2C, D)의 형태적인 특성과 포자(Fig. 2E, F)의 형태를 통해 분리한 균들이 Trichoderma 속과 Penicillium 속균으로 추정되었다. Trichoderma 속으로 추정되는 감염균의 분생포자의 크기는 직경 약 5 µm의 원형이었다(Fig. 2E). Penicillium 속의 균은 분생자경에 분생포자가 연쇄상으로 형성된 것을 관찰할 수 있었으며, 포자의 크기는 약 4 µm의 크기의 원형이었다(Fig. 2F). 이러한 형태적 동정의 특징은 Trichoderma 속균(Gezgin 등, 2023)과 Penicillium 속균(Visagie 등, 2014)에 대해 기술한 형태적 동정의 특징과 유사하였다.
분자생물학적 동정을 위해 ITS 영역의 염기서열을 확보하여 검정하였다. 그 결과 장흥에서 분리된 10개의 균주는 모두 Trichoderma pleuroticola (GenBank accession no=JQ040377)과 99.86%의 유사도를 보였다(Table 1). 순천의 목이버섯 농장의 오염배지에서 분리된 44개 균주들의 ITS 염기서열 검정 결과40개(90.9%) 균주가 Trichoderma pleuroticola (GenBank accession no=JQ040377)와 98.56-99.84%의 유사도를 보였다. 3개(6.8%) 균주는 Penicillium citrinum과 GenBank accession nos. OR346125, MT529486, MN78102로 각각 99.5%, 100%, 100%의 유사도를 보였다. 1개(2.3%) 균주는 Talaromyces wortmannii (GenBank accession no=MH858298)와 100%의 유사도를 보였다. 이 결과를 통해 목이버섯 농장 두 곳에서 분리된 균주 총 54개 중 Trichoderma 속이 50개(92.6%), Penicillium 속이 3개(5.6%), Talaromyces 속이 1개(1.8%)를 차지하여 Trichoderma 속이 우점하였다.
종 동정을 위해서는 ITS 영역 염기서열 이외에도 다양한 유전자 정보의 염기서열이 필요하다. Trichoderma 속은 ITS 영역 및 RPB2 유전자 염기서열 정보가 필요하다. 이와 더불어 더 정확한 동정을 위해 translation elongation factor 1, calmodulin (CaM) 유전자 염기서열 정보가 추가적으로 필요하다(Gezgin 등, 2023; Kamil 등, 2022; Kubicek 등, 2019; Maheshwary 등, 2022). 이 정보에 기초하여 대표 균주를 선발하기 위해, 이 실험에서 우점적으로 분리된 BR6-3과 10-1 균주의 RPB2 염기서열 정보를 확보한 뒤, NCBI 정보를 기반하여 Trichoderma sp. BR6-3 (GenBank accession number, PP129697)과 BR10-1 (PP129698) 균주가 모두 T. miyunense로 밝혀졌다. 이 두 균주를 공시 균주로 사용하기 위해 KACC에 기탁하여 KACC410667 (T. miyunense BR6-3)과 KACC410668 (T. miyunense BR10-1)로 명칭을 변경하여 이후 실험에 사용하였다.
또한 Penicillium 속의 동정을 위해서는 ITS, β-tubulin, CaM, RPB2 염기서열 정보가 추가적으로 이루어져야 한다(Visagie 등, 2014). 이 연구에서는 분리 균주의 나머지 barcoding 유전자에 대한 염기서열 정보가 확보되지 않았으므로, 앞으로 각 균주들에 대한 명칭은 Penicillium sp.로 사용하였다.
목이버섯 종균과 푸른곰팡이병균의 균사생장 비교.
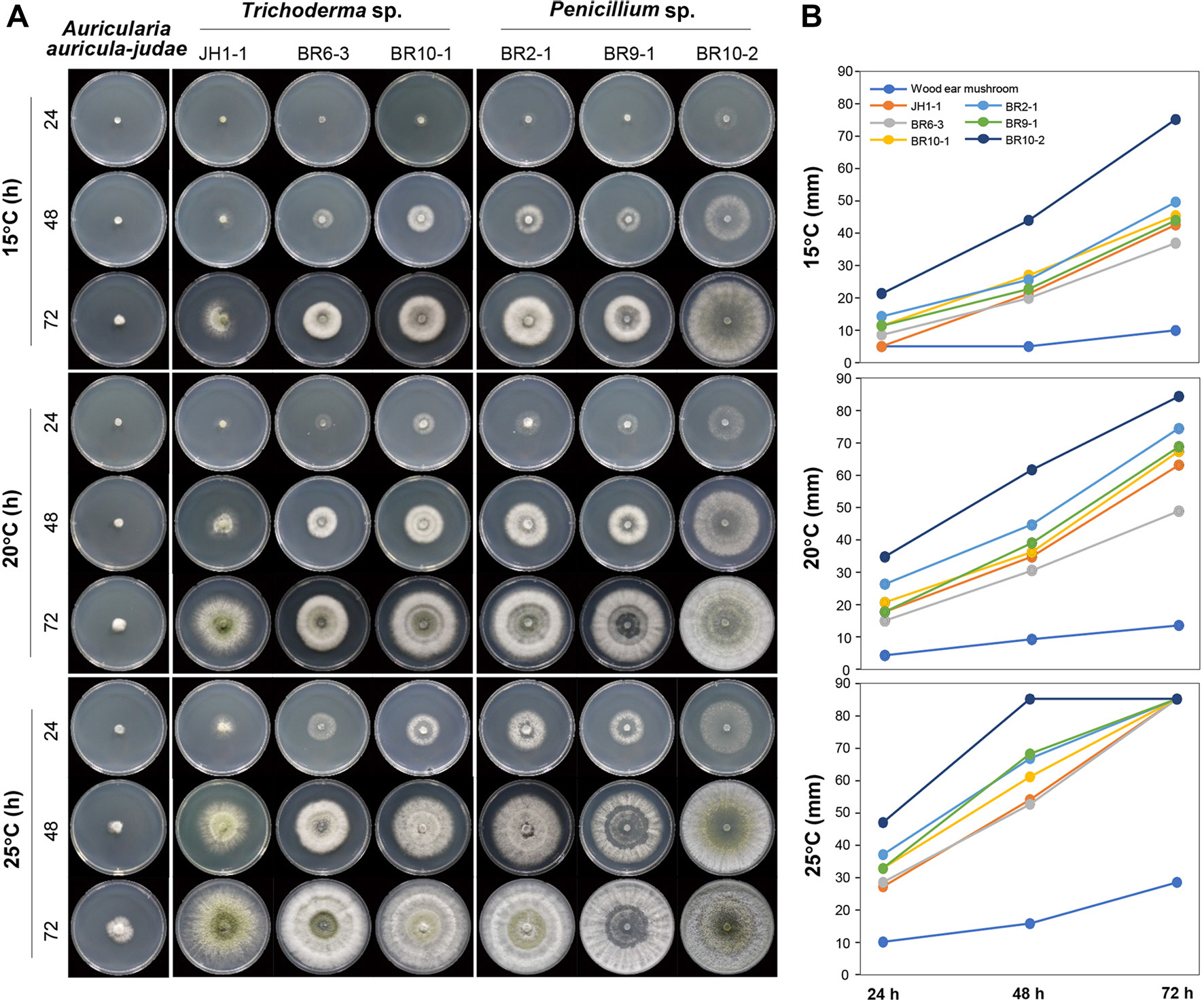
목이버섯 종균(A. auricula-judae)의 적정 생육 온도는 15-25° C이다. 이 온도 범위에서 목이버섯과 분리된 감염 곰팡이들의 상대적인 성장을 비교하고자 15-25° C 범위를 5° C 간격으로 나누어 균사 생장의 속도를 비교하였다. 실험을 위해 우점하는 Trichoderma 속균은 장흥에서 1균주(JH1-1)를 선발하였으며, 순천에서 2개 균주(KACC410667, KACC410668)를 선발하였다. Penicillium 속균의 경우는 총 3개의 균주(BR2-1, BR9-1, BR10-2)가 분리되어 모든 균주를 실험에 사용하였다. Talaromyces 속균은 분리 빈도가 너무 낮아 실험에서 배제하였다.
목이버섯 종균은 15-25° C 내의 모든 온도에서 느리게 성장하였다(Fig. 3). 그러나 Trichoderma 속균과 Penicillium 속균이 목이버섯 종균에 비해 모든 온도 범위에서 빠르게 자랐다. 특히, Penicillium sp. BR10-2 균주는 모든 온도 범위에서 PDA 배지에 접종한지 72시간 만에 9 cm 배지를 모두 점유하였다. 이 결과를 통해 목이버섯 종균이 성장하는 곳에 배지 감염 곰팡이균인 Trichoderma 속균과 Penicillium 속균이 감염할 경우 15°, 72시간 이후 경쟁 곰팡이가 4배(Trichoderma sp. JH1-1)에서 7.5배 (Penicillium sp. BR10-2) 빠르게 자랄 수 있으며, 온도가 올라갈수록 이러한 추세는 더욱 가속화될 것으로 판단되었다. 이 결과는 목이버섯 농장에 분리된 감염 곰팡이가 자라면 이들 곰팡이의 오염을 막기 어려울 것으로 생각된다(Fig. 3).
Fig. 3.
Comparison of the mycelial growth of wood ear mushrooms (Auricularia auricular-judae) with competitive fungi (three representatives each of Trichoderma sp. and Penicillium sp.) at different temperatures ranging from 15°C to 25°C. (A) Comparison of mycelial growth at a temperature range of 15-25°C with 5°C intervals up to 24 h (top), 48 h (middle), and 72 h (bottom). (B) Graphical display of the experimen-tal results.
목이버섯 푸른곰팡이병균에 대한 길항세균 선발.
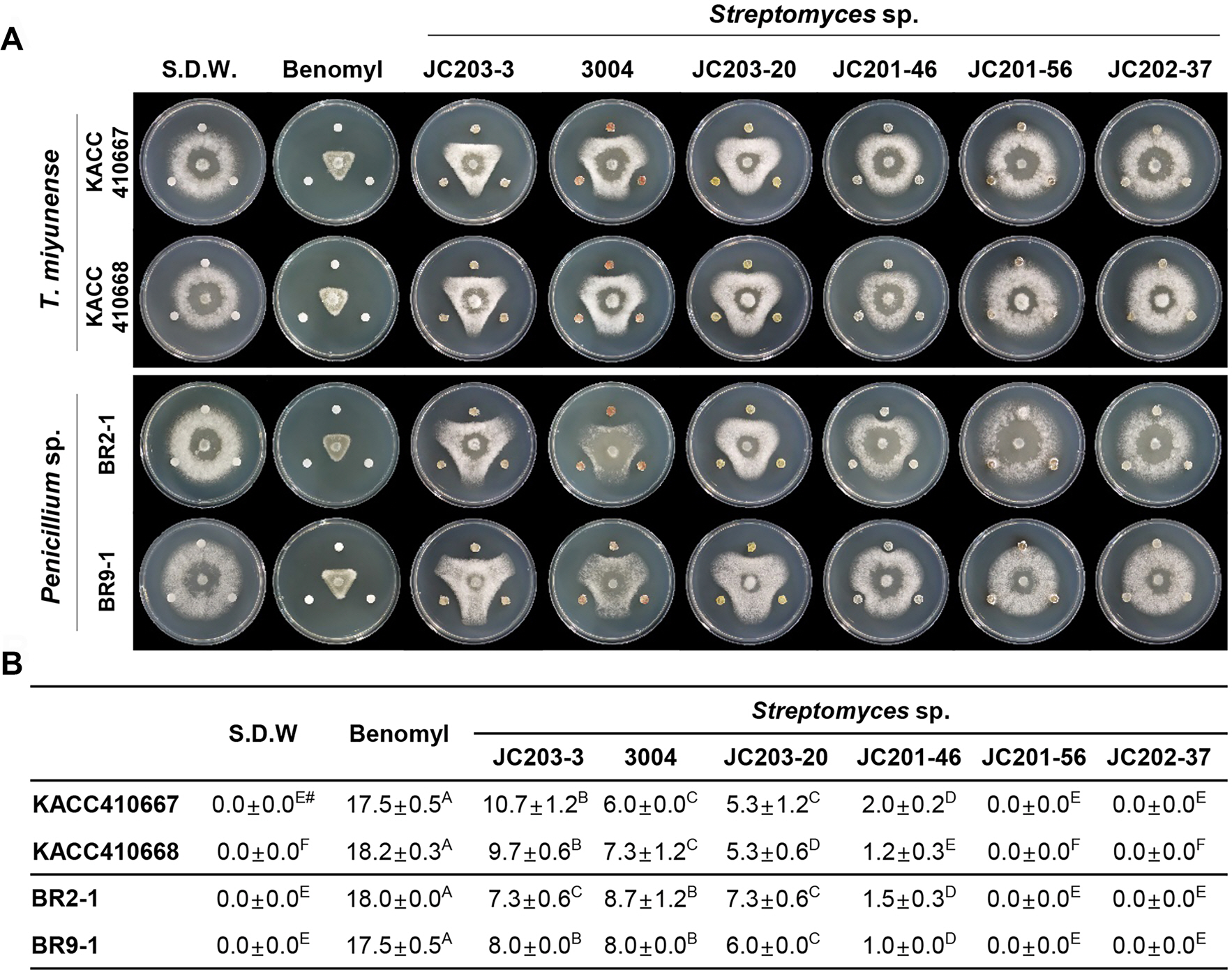
길항세균 선발을 위해 대표 균주로 2개의 T. miyunense (KACC410667와 KACC410668) 균주 및 2개의 Penicillium sp. (BR2-1과 BR9-1)를 대표 균주로 선택하여 실험에 사용하였다. 길항세균은 국립생물자원관에서 분양받은 6개의 Streptomyces sp. 균주(JS203-3, NIBRBAC00000300, JC203-20, JC201-46, JC201-56, JC202-37)를 검정 실험에 사용하였다. 음성 대조군으로는 멸균증류수, 양성 대조군으로는 농약 베노밀 수화제를 선정하였다. 베노밀 수화제는 Aspergillus nidulans의 이배체에서 핵의 불안정화를 유도하고, DNA 합성을 방해하여 궁극적으로는 핵 분열을 막아 세포분열을 저해하는 작용기작을 갖는 것으로 밝혀진 농약이다(Clemons 와 Sisler, 1971; Davidse, 1973; Hammerschlag과 Sisler, 1973). 이 실험에서 베노밀 수화제를 선정한 이유는 느타리버섯, 상황버섯, 표고버섯에 푸른곰팡이 제어 농약으로 등록되어 있기 때문이다(한국작물보호협회, https://www.koreacpa.org/ko).
Streptomyces sp. 3개 균주들(JC203-3, NIBRBAC00000300과 JC203-20)은 T. miyunense KACC410667과 KACC410668 균주들 및 Penicillium sp. BR2-1과 BR9-1 균주들에 대해 생장 억제 활성을 보였다. 그러나 나머지 Streptomyces sp. 3개 균주들(JC201-46, JC201-56 및 JC202-37)은 균사생장 억제 활성이 낮게 나타났다(Fig. 4). 특히, 공동배양 실험에서 베노밀 수화제가 경쟁 곰팡이들인 T. miyunense 및 Penicillium sp. 균주들의 균사생장을 효과적으로 제어하였는데(Fig. 4), 이 결과는 길항세균 중 가장 높은 균사생장 저해 억제능을 보였던 Streptomyces sp. JC201-46와도 통계학적으로 큰 차이를 나타냈다(Fig. 4B).
Fig. 4.
Screening of the medium-based antifungal activity of six Streptomyces sp. against two representatives each of Trichoderma miyunense and Penicillium spp. (A) Standard dual culture assay of the selected six Streptomyces spp. (JC203-3, NIBRBAC000003004, JC203-20, JC201-46, JC201-56, and JC202-37). Sterilized distilled water (S.D.W.) and benomyl were used as the negative and positive controls, respectively. (B) Effect of the dual-culture of six Streptomyces spp. on the growth of two representatives each of T. miyunense and Penicillium spp. Inhibition zone=mm. # Tukey's test was used to determine significance at the 95% probability level. The same letters in a column showed no significant difference.
Kunova 등(2016)은 Streptomyces 속 균주들의 길항성을 검정하기 위해 모든 실험을 동일하게 적용하기보다는 각 곰팡이의 균사생장을 속도를 고려하여 1) 빠르게 자라는 성장 그룹, 2) 중간 속도의 성장 그룹, 3) 느리게 성장하는 그룹으로 구분하여 Streptomyces 길항성을 검정해야 한다고 주장하였다. Streptomyces spp.의 2차 대사산물이 기중균사와 포자를 만드는 시기에 생성되기 때문이다(Kunova 등, 2016). 이 실험 결과를 적용하여, 우리는 앞선 1차 실험에서 수행한 일반적인 공동배양 실험을 수정하여 다른 공동배양법을 적용하였다.
목이버섯 배지 감염 곰팡이들(Trichoderma 종과 Penicillium 종)의 경우는 9 cm, PDA 배지에서 3일만에 균사가 생장하는 매우 빠르게 자라기 때문에(Fig. 3), 3개의 길항세균인 Streptomyces sp. JC203-3, NIBRBAC00000300과 JC203-20를 공동배양 실험 3일 전에 PDA 배지의 중앙에 접종하였다. 그 결과 Streptomyces sp. JC203-3 균주는 농약인 Benomyl 수화제보다도 통계학적으로 유의한 수준에서 모든 대상 곰팡이의 균사생장을 억제하였으며, Streptomyces sp. NIBRBAC00000300 균주의 경우도 Benomyl 수화제만큼의 균사생장 억제 효과를 보여 Streptomyces 균주들의 우선 접종에 따른 높은 길항 효과를 검정할 수 있었다(Fig. 5). 이 결과는 Streptomyces sp. JC203-3과 NIBRBAC00000300 균주들이 버섯 재배지에 발생하는 푸른곰팡이를 억제할 수 있는 잠재적 친환경제제로서의 가능성을 시사한다.
Fig. 5.
Medium-based antifungal activity of three selected Streptomyces spp. (JC203-3, NIBRBAC000003004, JC203-20) against two representatives each of Trichoderma miyunense and Penicillium spp. (A) Dual culture assay of three selected Streptomyces spp. Streptomyces spp. were inoculated on potato dextrose agar media 3 days prior to the dual-culture assay. (B) Effect of the dual-culture of three Streptomyces spp. on the growth of two representatives each of T. miyunense and Penicillium spp. Inhibition zone=mm. # Tukey's test was used to determine significance at the 95% probability level. The same letters in a column showed no significant difference. S.D.W., sterilized distilled water.

목이버섯을 포함하는 다양한 식용 버섯들은 담자균에 속하는 곰팡이로 우리나라 2023년을 기준으로 542 ha 재배면적에 142,352톤의 생산량을 자랑하는 주요 식용 식품 중 하나이다(Statistics Korea, 2023). 버섯 재배지에서 이용되는 배지 재배는 푸른곰팡이로 알려진 Trichoderma 속 및 Penicillium 속과 같이 균사의 생장이 폭발적으로 빠른 곰팡이에 감염될 경우 버섯 생산에 수확량 손실을 유발한다(Houbraken 등, 2019; Kim 등, 2019; Kosanović 등, 2015; Lee 등, 2017; O'Brien 등, 2014; O'Brien 등, 2017).
한국에서는 표고버섯(Kim 등, 2019), 느타리버섯(Park 등, 2006), 느티만가닥버섯(Kim 등, 2020)에 발생하는 푸른곰팡이에 대한 균주의 분리 및 동정에 관한 연구가 보고되었다. 특히 표고버섯의 푸른곰팡이의 억제를 위해서는 항진균성 펩티드를 적용하는 연구도 보고되었다(Lee 등, 2017). 따라서 이 연구의 목적은 아직까지 보고된 바 없는 목이버섯 배지에 발생하는 푸른곰팡이에 관한 우점종을 밝혀내고, 친환경제제의 선발을 목적으로 하였다. 특히 이 연구에서는 Kunova 등(2016)과 마찬가지로 Streptomyces 균주들이 포자를 형성하는 시기를 고려하여 3일 전에 우선 접종하는 방식을 적용하여 판매되는 농약보다도 높은 길항 효과를 보이는 균주들을 선발할 수 있었다. 따라서 친환경제제를 개발하더라도 포자가 발생하기 시작하는 시점을 고려해야 할 것이다.
농약허용물질목록관리제도(PLS)가 2019년부터 시행됨에 따라, 농약사용 기준에 대한 잔류 허용 기준에 대한 경각심이 높아지고 있다. 버섯의 특성상 세척하지 않고 소비되는 약제 처리는 잔류 농약의 위험성을 동반하게 되므로 대체 가능한 친환경 제제의 필요성은 점차적으로 요구되고 있다. Streptomyces 속과 같은 친환경제제의 선발 및 개발은 앞으로의 농민들의 고부가가치를 올릴 수 있는 대체적인 약제라 할 수 있다. 그러나 아직까지 선발된 균주들의 항생제 생산 여부나 혹은 또 다른 식품 적용에 대한 안전성 검정이 이루어져야 할 것이다. 그러나 이 연구에서 선발된 Streptomyces 종들은 버섯 재배에 위협이 되는 Trichoderma 속균들과 Penicillium 속균들에 대한 잠재적 대체재로의 가능성이 높다. 따라서 농약 대체재로서의 잠재력을 평가하기 위해 추가적인 연구가 필요하다.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






