



서 론
경제협력개발기구(Organization for Economic Cooperation and Development)에 따르면 안정적이고 지속가능한 식량보급을 위하여 현재 대비 1.5배 이상의 작물의 생산성을 요구하고 있다(Jägermeyr 등, 2020; Xu 등, 2019). 2050년까지 세계 인구가 약 97억명에 이르고 기후와 관련된 작물 생산성이 감소할 것으로 예상되어져 안정적이고 지속가능한 농업은 식량안보 문제로 인식되고 있다(Chen 등, 2016; Lau 등, 2017; Rodriguez 등, 2019). 이러한 관점들은 인류가 필요로 하는 농업의 녹색 혁명을 강화하고 phytobiome 개념을 도입시켰다. Phytobiome은 미생물 인자와 같이 식물을 보호하고, 생산성을 높이기 위한 주변 인자가 상호 유익한 방향으로 작용하며 식물재배와 관련된 모든 인자를 의미한다(Xiong 등, 2021).
그 중 식물 관련 미생물은 토양부터 식물의 화기까지 식물의 모든 조직을 미생물의 생존 공간으로 사용하고 있으며 각각, 근권(rhizosphere), 내권(endosphere), 엽권(phyllosphere) 및 종자생물군계(seedbiome)로 분류된다(Bintarti 등, 2022; Hannula 등, 2019; Kim 등, 2019a; Lu 등, 2018). 또한 지난 수십년 동안 연구되어 온 plant growth-promoting rhizobacteria (PGPR) 관련 미생물은 생물적 ‧ 비생물적 스트레스로 인한 피해를 일정부분 경감시킬 수 있는 것으로 알려져 있다(Berkelmann 등, 2020). PGPR 미생물의 관련 기작으로 IAA (indole-3-acetic acid) 생산, 질소 고정, siderophore 생산 등 다양한 경로를 통한 식물 생장 촉진 능력을 가지고 있으며 대표적인 미생물 군으로 Bacillus, Pseudomonas 그리고 Streptomyces가 있으며 이들은 식물의 생장을 향상시키는 직간접 메커니즘을 가진 것으로 알려져 있다(Andleeb 등, 2022; Bokhari 등, 2019; Fidan 과 Zhan, 2019; Herrera-Quiterio 등, 2020; Oviedo-Pereira 등, 2022).
그 중 Streptomyces 속은 식물의 근권, 엽권 그리고 내권에서 군집화가 가능한 것이 확인되었다(Lindow와 Brandl 2003; Redford 등, 2010). 또한 1940년 actinomycin 발견 이후, 생물학적 활성을 가진 항생제 및 유용 2차 대사산물의 확보에 핵심 미생물로 인지되고 있다(Bentley 등, 2002; Deshpande 등, 1988; Lee 등, 2020). 현재까지 알려진 항생제 총 70-80%의 항생제가 Streptomyces속에서 분리되어 보고되었다(Harir 등, 2018). Streptomyces 이차대사산물과 관련된 기존의 연구에 따르면, Streptomyces의 생활상 단계 중 기균사(aerial hypha)로부터 포자가 형성되는 단계에서 이전 단계에서 생성되지 않았던 2차 대사산물의 전환이 이루어지므로 포자의 형성은 방선균의 항생물질 생산에 있어 중요한 과정으로 알려져 있다(Yagüe 등, 2012).
선행연구에서 확보된 실물 자원 Streptomyces globisporus SP6C4는 딸기 시들음병과 화기에서 발생하는 잿빛곰팡이병에 대하여 유용미생물로 잘 특성화되었으며, 이는 대상 식물병 발생 억제에 SP6C4균주가 생산하는 내열성 화합물과 작용이 관여함이 밝혀졌다(Cha 등, 2016; Kim 등, 2021). S. globisporus SP6C4는 2가지의 신규 lantipeptide를 포자형성 단계에서 생산하며, 이는 conprimycin과 grisin으로 진균 세포벽 생합성 및 RNA 중합효소 활성을 억제하였다(Cha 등, 2016; Kim 등, 2019b). 또한 Streptomyces sp. S8은 잔디 재배지 및 사용지에서 발견되는 large patch(갈색마름병)를 발생시키는 병원균 Rhizoctonia solani AG2-2의 생장을 억제할 수 있는 탁월한 길항 특성을 가지고 있는 것으로 보고되었다(Jeon 등, 2019). Streptomyces sp. S8의 genome sequencing 정보를 바탕으로 CRISPR/Cas9 system을 통해 valinomycin 생합성 유전자 삭제 돌연변이를 제작을 통하여, 해당 돌연변이는 large patch 병원균에 대한 항진균 활성이 상실되는 것을 확인하였다(Jeon 등, 2019).
식물병의 발생을 효과적으로 억제하는 기능형 유용미생물은 실용화를 위하여 대량 배양 공정이 반드시 필요하다(Park 등, 2018, 2019). 하지만 미생물의 순수배양은 주로 고가의 실험용 배지가 사용되므로 농업현장에서 이를 활용하기에는 아직 많은 한계점이 존재한다(Park 등, 2018). 또한 유용미생물 제제로 활성을 나타내며 시판되기까지는 많은 공정이 필요하며, 그중 방선균은 배양에 있어 까다로운 특성이 있으므로 현재까지 배양 조건을 확립하는 것이 우선되어야 한다(Jung 등, 2011). 농업미생물의 대량배양시, 일반적으로 배양대상 미생물의 생장이 촉진되는 탄소원, 질소원에 대한 선정이 가장 중요하다. 또한 배지의 산도, 배양온도, 배양기의 회전수 등이 고려 대상이다(Lee 등, 2021). 본 연구에서는, 항진균 물질이 구명된 2종의 유용 방선균을 활용하여 개별 미생물 특이적으로 생장과 포자형성에 관여하는 물질을 구명하고자 하였다. 기 선정된 유용방선균 2종의 대량생산공정에 필요한 탄소원과 질소원을 규명하는 기초연구를 바탕으로 생물학적 방제제 개발에 대한 기초 자료를 마련하고자 한다.
재료 및 방법
PGPR 효소 활성 검증
Streptomyces sp. S8는 2017년 잔디 재배지의 근권에서 분리되었으며, 지하부 진균병원균에 대한 우수한 항진균력을 가지고 있다(Jeon 등, 2019). S. globisporus SP6C4 균주는 딸기 꽃에서 분리된 방선균으로 지상부 진균병인 잿빛곰팡이병, 꽃곰팡이병의 발생을 효과적으로 억제하는 특성이 보고되었다(Kim 등, 2019a). Streptomyces sp. S8과 S. globisporus SP6C4의 포자 현탁액 제조를 위해 MS media (mannitol 20 g, soya flour 20 g, agar 20 g per liter)에 도말하여 28°C 배양기에서 7일간 배양하여 포자를 형성하였다. 배양이 완료된 plate에 멸균솜과 1 ml 멸균수를 분주하여 포자를 멸균수로 침출시켰으며, 10-mL syringe를 통하여 filtering한 뒤 회수된 포자의 농도를 분광광도계(Synergy H1 Hybrid Multi-Mode microplate reader, BioTek, Winooski, VT, USA) OD600nm 로 0.4 (106 cfu/ml)로 조정하였다.
6종류의 PGPR 효소 활성을 검증하였으며, 진균 세포벽 와해 능력을 알 수 있는 chitinase와 cellulose 효소 활성과 철 이온 흡수 능력, 인산 가용화 효소, 질소 고정능력을 검증하였다. Chitinase 효소 활성을 검증하기 위해 chitinolytic reaction media를 제조하고자 5 g of chitin, 5 g of yeast extract, 0.7 g of KH2 PO4, 0.5 g of K2 HPO4, 0.3 g of MgSO4, 0.1 g of FeSO4, 0.1 g of NaCl를 1 liter 증류수에 넣고 pH 6.5-7.0로 조정한 다음 18 g of agar를 첨가하였고 proteinase 형성 능력 검증을 위해 skim milk (10 g skim milk, 18 g agar per liter) 배지를 사용하였다. 철 이온 흡수 능력을 검증하기 위해 CAS agar (solution 1: 0.06 g of CAS, 50 ml of ddH2 O; solution 2: 2.7 mg FeCl3-6H2 O, 10 ml 10 mM HCl; solution 3: 73 mg hexadecyltrimethylammonium bromide, 40 ml ddH2 O; Blu dye: solution 1: 50 ml, solution 2: 9 ml, solution 3: 40 ml; mixture solution: MM9 salt solution 500 ml [15 g KH2 PO4, 25 g NaCl, 50 g NH4 Cl, 500 ml of ddH2 O]; 20% glucose stock 100 ml, NaOH solution [25 g NaOH per 150 ml ddH2 O]; casamino acid solution [casamino acid 3 g per 27 ml ddH2 O]; CAS agar: 100 ml MM9 salt solution, 32.24 g PIPES, 15 g agar, 30 ml casamino acid solution, 10 ml 20% glucose solution, 100 ml blue dye)를 사용하였다. 인산 가용 능력을 검증하기 위해 Pikovaskaya's 배지(10 g glucose, 0.5 g (NH4)2 SO4, 0.2 g KCl, 0.1 g MgSO4, 0.002 g MnSO4, 0.002 g FeSO4, 0.5 g yeast extract, 5 g Ca3(PO4), 20 g agar per liter, pH 6.8-7.0)를 사용하였다(Kim 등, 2019c). 질소고정 능력을 검증하기 위해 액상의 NFb 배지(5 g malic acid, 0.6 g K2 HPO4, 0.4 g KH2 PO4, 0.01 g MnSO4, 0.05 g MgSO4, 0.02 g NaCl, 0.002 g Na2 MoO4, 2 ml of bromothymol blue [0.5% in alcohol], 1.75 g of agar per liter, pH 6.6-7.0.)를 사용하였다(Kim 등, 2019c). IAA production assay를 검증하고자 Salkowski reagent method (35% of HClO4 and 0.5 M of FeCl3)를 사용하였다(Patel과 Parmar, 2013).
해당 유용미생물 2종을 각각 tryptophan 함유된 배지와 함유되지 않은 PDK broth (10 g of potato dextrose broth, 2 g of Bacto peptone per liter)에 접종하여 30°C에서 14일간 배양한 다음 5,000 rpm 원심분리기로 10분간 반응시킨 후 1 ml의 상등액을 test tubes에 첨가하였다. 이 후 Salkowski reagent와 2:1 비율로 혼합 후 25분이 경과하였을 때 색상의 변화를 관찰하였다(Patel과 Parmar, 2013). Chitinase, siderophore, 질소고정능력, 그리고 protease 형성 능력은 28°C에서 배양하면서 24시간 간격으로 3일이 경과한 시점까지 확인하였으며, 단 질소고정 능력은 동일한 온도에서 일주일간 반응을 관찰하였다.
pH에 따른 생장 조건 및 포자 형성 능력 검증
생물학적 스트레스 저감 및 군집화 능력이 검증된 방선균 2종(Streptomyces sp. S8 and S. globisporus SP6C4) 균주의 생장 특성을 확인하고자 pH 2, 5, 7, 7.2, 10 조건에서 생장 속도를 분광광도계(Synergy H1 Hybrid Multi-Mode microplate reader, BioTek) OD600nm에서 0.2로 측정하였으며 포자 형성 능력과 배지 조건별로 ISP 2 media (yeast malt agar: yeast extract 4 g, malt extract 20 g, dextrose 4g, agar 20 g per liter), ISP 4 media (inorganic salt starch agar: CaCo3 2 g, K2 HPO3 1 g, MgSO4‧7H2 O 1 g, NaCl 1 g, (NH4)2 SO4 2 g, Trace salt solution [FeSO3‧7H2 O 0.1 g, MnCl2‧4 H2 O 0.1 g, ZnSO4‧7H2 O 0.1 g per 100 ml], ddH2 O 500-ml and adjusted pH 7.4), ISP 6 media (peptone yeast extract iron agar: peptone 15 g, proteose peptone 5 g, ferric ammonium citrate 0.5 g, dipotassium phosphate 1 g, sodium thiosulfate 0.08 g, yeast extract 1 g, agar 20 g per liter, adjust to pH 7.2), ISP 7 media (tyrosine agar: L-asparagine 1 g, L-tyrosine 0.5 g, dipotassium phosphate 0.5 g, magnesium sulphate·7H2 O 0.5 g, sodium chloride 0.5 g, sodium salt solution [ferrous sulphate·7H2 O 1.36 mg, copper chloride 2H2 O 0.027 mg, co-balt chloride 6H2 O 0.04 mg, sodium molybdate 2H2 O 0.025 mg, zinc chloride 0.02 mg, boric acid 2.85 mg, manganse chloride·4H2 O 1.8 mg, sodium tartarate 1.77 mg] agar 20 g per liter, adjusted pH 7.4). 그리고 MS (mannitol soya agar: mannitol 20 g, soya flour 20 g, agar 20 g per liter)에서 24시간 간격으로 72시간 동안 배양을 통해 확인하였다. 포자형성은 각각의 배지에서 방선균의 기균사 형성과 포자형성 정도는 육안으로 확인하였다.
Phenotype assay 를 이용한 질소원 선호도 검증
단일 균주를 대상으로 생장 시간을 단축시키기 위해 생장을 촉진할 수 있는 질소원을 선별하고자 PM3B plate Biolog plate (Biolog, Hayward, CA, USA)를 사용하였다. 각 plate에는 OD600에서 0.2로 조정된 방선균 포자 현탁액 10 µl를 분주한 후 28°C에서 3일간 배양하였다. 배양이 완료된 plate에 Redox dye mix A를 각 well에 10 µl 분주한 후 37°C에서 30분 간격으로 색의 변화를 관찰하였고 이후 선발된 질소원을 0.2% (final concentration)으로 질소원이 결핍된 basal media (KCl 0.05%, KH2 PO4 0.1%, MgSO4·7H2 O 0.05% and FeSO4·7H2 O 0.001%)에 농도를 5 μ M, 10 μ M, 50 μ M, 5 mM, 10 mM, 50 mM로 다르게 첨가하여 균주의 생장 속도에 영향을 미치는지 검증하였다(Ripa 등, 2009).
탄소원 및 질소원 첨가에 따른 포자 형성 능력 검증
포자 형성 능력 검증에서 확보된 탄소원과 Biolog 3B plate를 통해 생장을 조절하는 것으로 선발된 질소원을 이용하여 액상 배양 시 포자 형성 능력을 검증하였다. 탄소원과 질소원을 각각 basal media (KCl 0.05%, KH2 PO4 0.1%, MgSO4·7H2 O 0.05%, FeSO4·7H2 O 0.001%)에 final concentration 0.2%로 첨가하였으며, 모든 탄소원과 질소원은 2% stock으로 제조하여 0.2 μ m syringe filter 후 사용하였다. 이후 OD600 nm 0.2로 조정된 포자 현탁액 10 µl를 1 ml의 basal media에 접종하여 30°C에서 진탕배양으로 3일간 배양 후 현미경(BX53 microscope, Olympus, Tokyo, Japan) 1,000배 배율에서 포자의 형성 정도를 관찰하였다.
결과 및 고찰
Streptomyces 성장은 숙주 식물의 공급에서 관찰되는 것처럼 자연 조건에서 특정 영양 공급원에 의해 향상될 수 있다(Costa 등, 2002; Reese 등, 2018). 식물에서 특정 탄소원과 질소원 영양소는 미생물의 성장과 밀도를 지원하고 조절할 수 있으며, 세포 성장을 향상시킬 뿐만 아니라 미생물과 숙주의 기능적 상호 작용에 크게 기여한다(Du 등, 2021; Ducray 등, 2019). 이들 화합물은 항생 기능을 가진 생체 활성 화합물 및 단백질의 생산에 조절자로 역할을 수행하거나 활용된다고 보고되어 있다(Costa 등, 2002; Rico와 Preston, 2008; Voelker 와 Altaba, 2001). 그 예시로 Kim 등(2021)은 L-glutamic acid 가 지상 및 지하 식물 조직 모두에서 핵심 박테리아 균주의 개체군을 재건하고, 미생물 군집 구조를 재구성하며 높은 미생물 다양성을 유지하여 최종적으로 식물 병원체의 공격을 예방할 수 있다고 보고하였다.
방선균은 균주 특이성이 매우 높은 특성을 지니고 있으며 2차 대사산물의 형성이 중요하다(Mostafa, 1979). 또한 생태계 내 생존을 위해 자체 생산하는 2차 대사 물질 분해 능력을 지니고 있으므로, 1차 대사를 충분히 진행한 상태에서 2차 대사로 전환율을 단시간에 높일 수 있도록 유도하여야 한다(Padilla 등, 1991). 이에 기초 배양 조건 단계로서 방선균의 영양요구성에 따라 선발된 탄소원, 질소원 혼합 조건을 이용하여 포자 형성 능력과 생장 속도 측정을 통하여 균주 생장 속도에 있어 우수하다고 보여지는 배지 혼합형을 이용하여 선발하고자 하였다. 또한 마지막 대량 배양 단계로서 배양액을 접종원으로 하여 대량배양기에서 조절할 수 있는 적합 pH 조건을 선발하고자 하여 방선균 최적합 배양 조건을 확보하고자 하였으며, 균주의 특성 구명을 위해 6종류의 PGPR 활성 능력을 함께 검증하였다.
PGPR 효소 활성 검증 결과
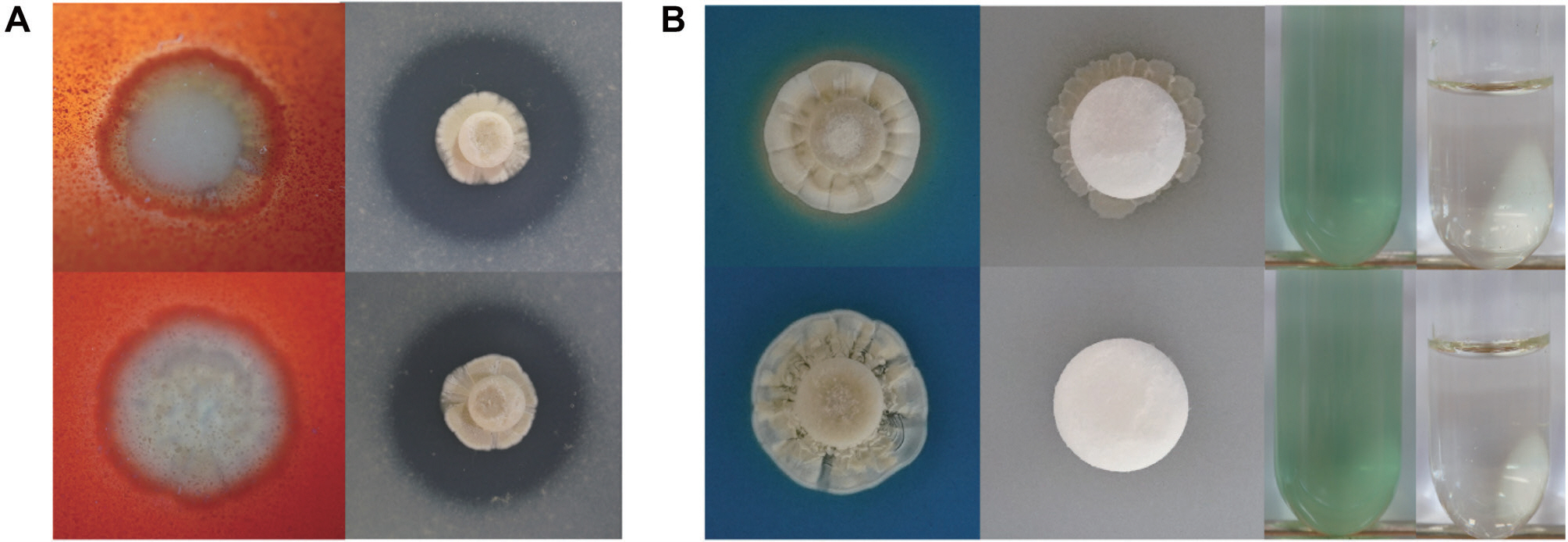
실물자원으로 기 확보된 2종의 방선균인 Streptomyces sp. S8과 Streptomyces globisporus SP6C4은 각각 서로 다른 기주에서 분리되었지만 항진균 활성이 높다는 공통적인 특징을 지니고 있었다(Cha 등, 2016; Jeon 등, 2019; Kim 등, 2019a). 이에 본 연구에서 두 균주의 특성을 구명하고자 항진균 활성을 나타내는 2종인 chitinase enzyme production과 protease enzyme production을 포함하여 다양한 효소 활성을 검증할 수 있는 siderophore production, phosphate solubilization activity, nitrogenase enzyme production, IAA production을 확인하였다. 그 결과 S8 균주는 chitinase, protease, siderophore, phosphate solubilization에서 모두 활성을 나타냈으며, IAA 효소 생성 능력과 질소고정능력은 확인되지 않았다(Table 1, Fig. 1). 하지만 SP6C4는 protease를 제외한 모든 효소 활성 능력이 없는 것으로 확인되었다(Table 1, Fig. 1). 총 6종류의 PGPR 효소 능력 검증 결과 S8 균주가 SP6C4 보다 다양한 활성을 나타내는 것으로 확인되었다.
Fig. 1.
Plant growth-promoting rhizobacteria activity of Streptomyces sp. S8 and S. globisporus SP6C4. The spore stock solution (10 µl) was added to each enzyme assay medium that contained an 8 mm diameter filter paper disk. (A) Chitinase enzyme production (left), Protease enzyme production (right). (B) Siderophore production, phosphate-solubilization activity, nitrogenase enzyme production, and indole-3-acetic acid production. The upper panel in each figure showed S8 and the lower panel presented SP6C4 strain.
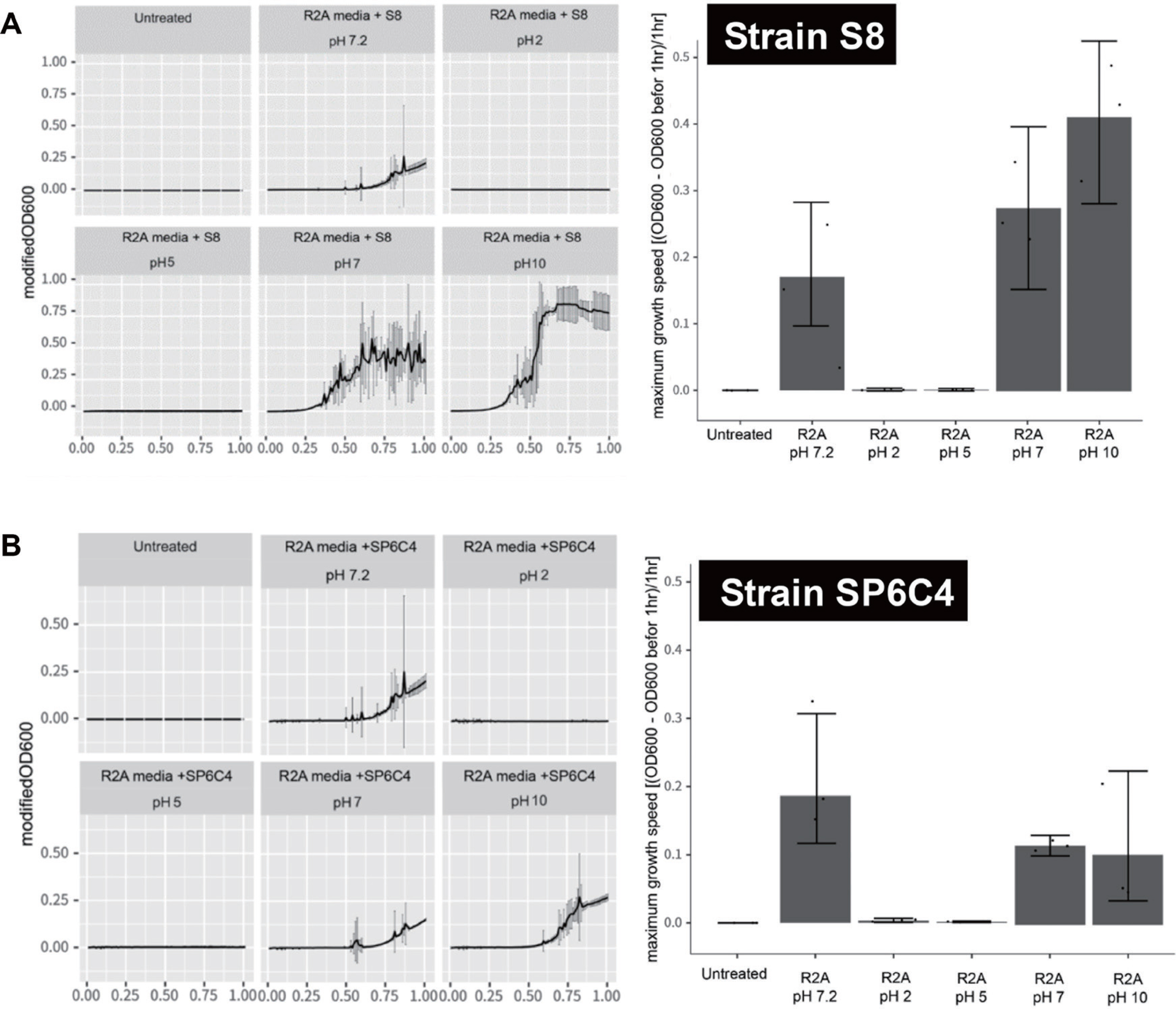
pH에 따른 생장 조건 및 포자 형성 능력 검증 결과
대량 배양 시 초기 생장률을 촉진할 수 있는 적합 pH를 검증하고자 산성 조건인 pH 2와 pH 5, 중성 조건인 pH 7와 pH 7.2, 그리고 염 기 조건인 pH 10을 이용하여 방선균의 생장률과 비생장률(기울기를 이용하여 시간당 최대 생장률) 측정하였다. 그 결과 S8균주는 pH 7과 pH 10에서 생장이 가능함을 보였으며 염기 조건인 pH 10에서 가장 빠른 생장률을 확인할 수 있었고, SP6C4는 중성 조건인 pH 7, pH 7.2 그리고 염기 조건인 pH 10에서 생장률을 나타냈지만 중성인 pH 7.2에서 가장 높은 생장률을 나타내는 것으로 확인되었다(Fig. 2).
Fig. 2.
Variation with pH in bacterial growth. The spore stock (10 µl) of the suspension was added to each well of a 96-well plate. The plates were incubated, and the optical density was measured with orbital shaking using a Synergy H1 Hybrid Multi-Mode microplate reader for a period of 24 hr at a temperature of 28°C. (A) S8 strain. (B) SP6C4 strain. The left panel showed bacterial growth in each pH condition. The right panel presented the maximum growth speed of the strain.
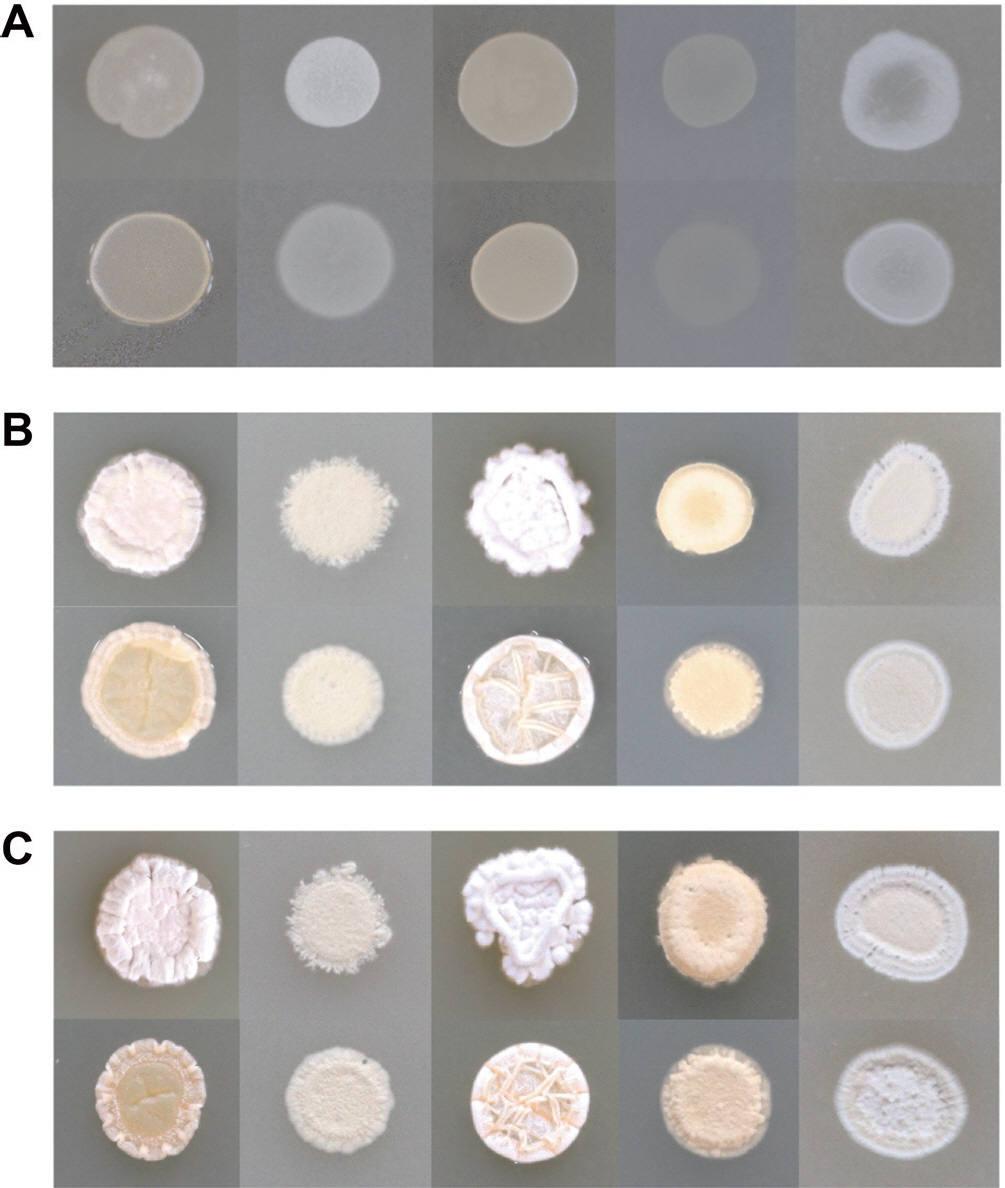
포자 형성 능력 검증에 사용한 5종류의 배지에서 24시간 간격으로 72시간까지 확인한 결과 S8과 SP6C4 모두 24시간에서 ISP7에서 가장 약한 포자 형성 능력을 나타냈었다(Fig. 3A). 48시간이 경과하였을 때 S8은 ISP2와 ISP6 그리고 MS에서 빠른 포자 형성 능력이 확인되었으며 이는 SP6C4도 유사한 결과로 확인되었다(Fig. 3B). 72시간 경과 후 5가지 배지에서 모두 포자 형성이 확인되었으며 S8이 SP6C4보다 초기 포자 형성 능력이 우수하며, 두 균주 모두 공통적으로 yeast와 mannitol을 주 탄 소원으로 함유된 배지에서 포자 형성이 유도되는 것으로 확인되었다(Fig. 3C).
Fig. 3.
The emergence of phenotypic in colonies of Streptomyces sp. S8 and S. globisporus SP6C4. Five different media, ISP2, ISP4, ISP6, ISP7, and MS (from left to right) were tested. The plates were incubated at 28°C for 72 hr, and observations were made at three different time points: (A) 24 hr after incubation, (B) 48 hr after incubation, and (C) 72 hr after incubation. The upper panel showed S8 and the lower panel presented SP6C4 strain.
Phenotype assay를 이용한 질소원 선호도 검증 결과
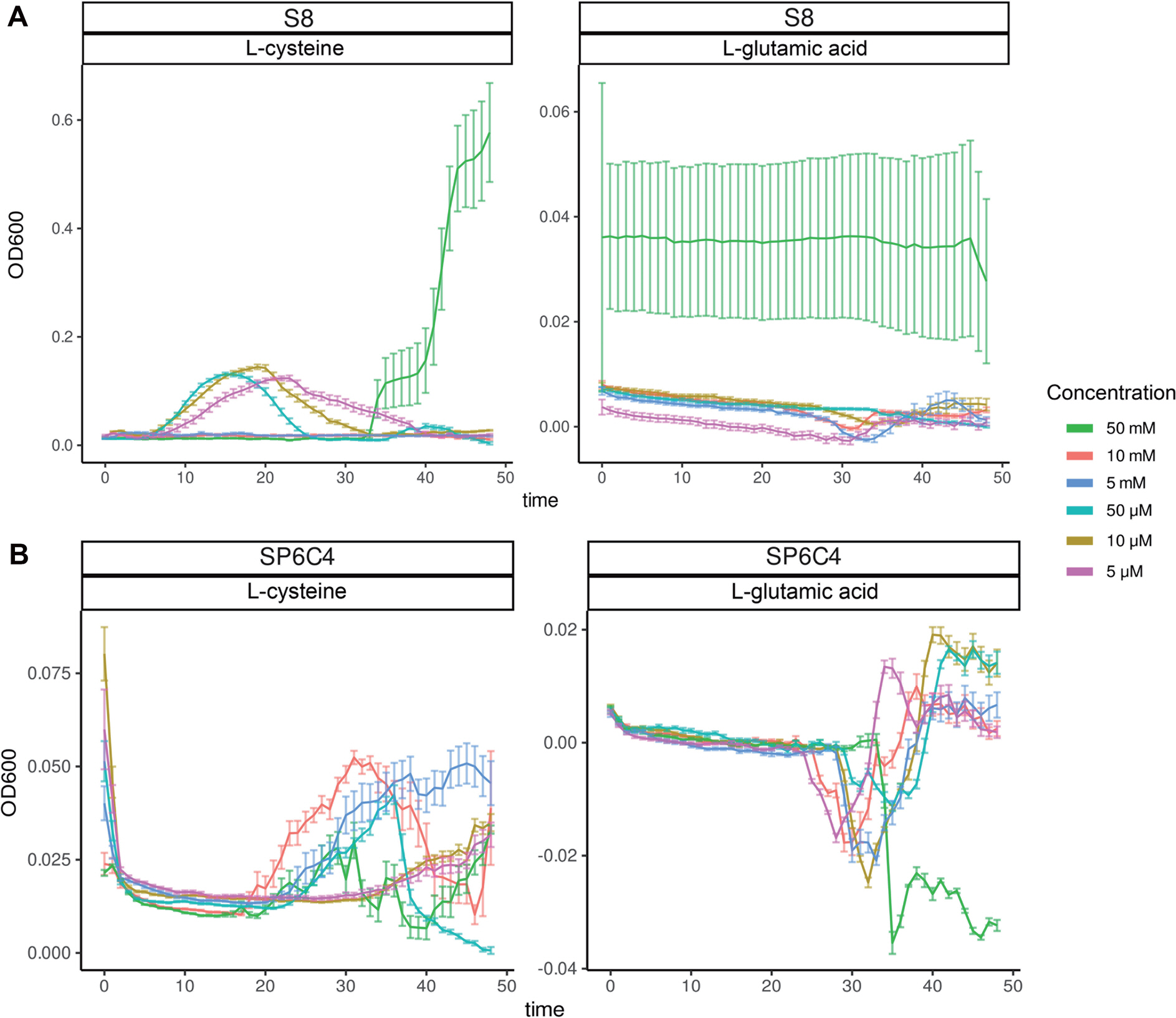
탄소원으로서 2균주 모두 적용 가능한 yeast와 mannitol을 선발한 다음 질소원을 선발하고자 Biolog 3B plate를 실험에 사용하였다. 그 결과 S8은 단일 질소원으로 ammonia와 L-cysteine 에서 생장이 이루어진 것을 확인하였으며 SP6C4는 L-cysteine, L-glutamic acid, L-proline으로 확인되었다(Fig. 4). 각각 선발된 질소원을 농도에 따라 사용하여 생장률을 확인한 결과 S8은 L-cysteine이 50 mM 농도로 첨가되었을 때, 그리고 SP6C4는 L-glutamic acid 첨가되었을 때 최고 농도인 50 mM을 제외하고 생장이 모두 촉진되는 것을 확인하였다(Fig. 5).
Fig. 4.
Biolog phenotype array for nitrogen utilization by PM3B plate. The spore stock was inoculated onto PM3B plates. A re-dox dye mix A was added to the plate, and the plate was further incubated at 37°C. Numbers (1-12) and alphabet (A-H) repre-sented well position and each position contained different nutrient sources.
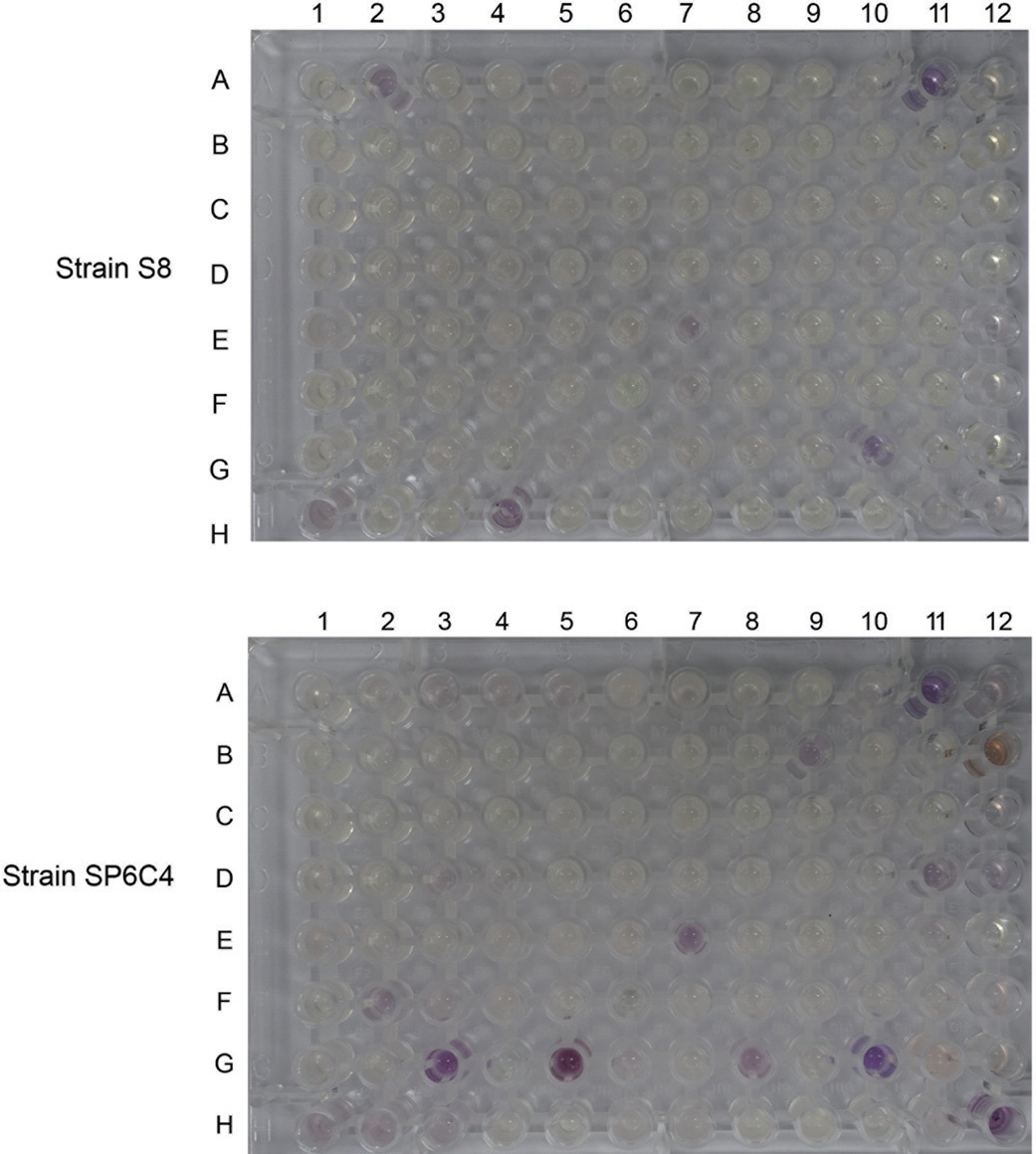
Fig. 5.
Effect of differences in nitrogen sources to bacterial growth. The nitrogen stock solution was added to basal media, followed by the addition of bacterial spore stock standardized to an optical density of 0.2 at 600 nm. The mixture was then incubated with shaking at 28°C for a duration of 50 hr (x-axis).
탄소원 및 질소원 첨가에 따른 포자 형성 능력 검증 결과
각각에 선발된 탄소원 2종과 질소원 3종을 포함하여 방선균 포자 형성과 2차 대사산물 형성 과정에 관여한다 고 보고된 L-asparagine, L-valine, peptone과 starch를 이용하여 액상 배양 조건에서의 포자 형성 능력을 검증하였다(Kalaiyarasi 등, 2020; Mostafa, 1979). 그 결과 S8는 탄소원 중 starch와 mannitol에서 포자 형성이 이루어지지 않았으며 aggregation되는 것이 관찰되었고, yeast extract에서 포자 형성이 확인되었다(Fig. 6). 반면 질소원 6종에 대해서는 모두 왕성한 포자 형성이 확인되었다(Table 2, Fig. 6). 다른 방선균인 SP6C4에서는 탄소원 중 yeast extract에서만 포자 형성이 이루어졌으며 질소원의 경우 L-glutamic acid과 L-proline에서만 포자 형성이 관찰되었다(Fig. 7). 특이적으로 peptone이 첨가된 경우 SP6C4에서 기균사가 왕성하게 형성된 것을 확인하였다(Table 2, Fig. 7).
Fig. 6.
Spore chain formation with different carbon and nitrogen sources by Streptomyces sp. S8. Six different types of nitrogen sources and three types of carbon sources were added to basal media at a final concentration of 0.2%. The sporulation was visualized under a 1,000× microscope (scale bars=10 μm). (A) L-asparagine. (B) L-cysteine. (C) L-glutamic acid. (D) L-proline. (E) L-valine. (F) Peptone. (G) Starch. (H) Mannitol. (I) Yeast extract.

Fig. 7.
Spore chain formation with different carbon and nitrogen sources by Streptomyces globisporus SP6C4. Basal media was supple-mented with six different nitrogen sources and three different carbon sources, each at a final concentration of 0.2%. The sporulation was observed under a 1,000× microscope (scale bars=10 μm). (A) L-asparagine. (B) L-cysteine. (C) L-glutamic acid. (D) L-proline. (E) L-valine. (F) Peptone. (G) Starch. (H) Mannitol. (I) Yeast extract.
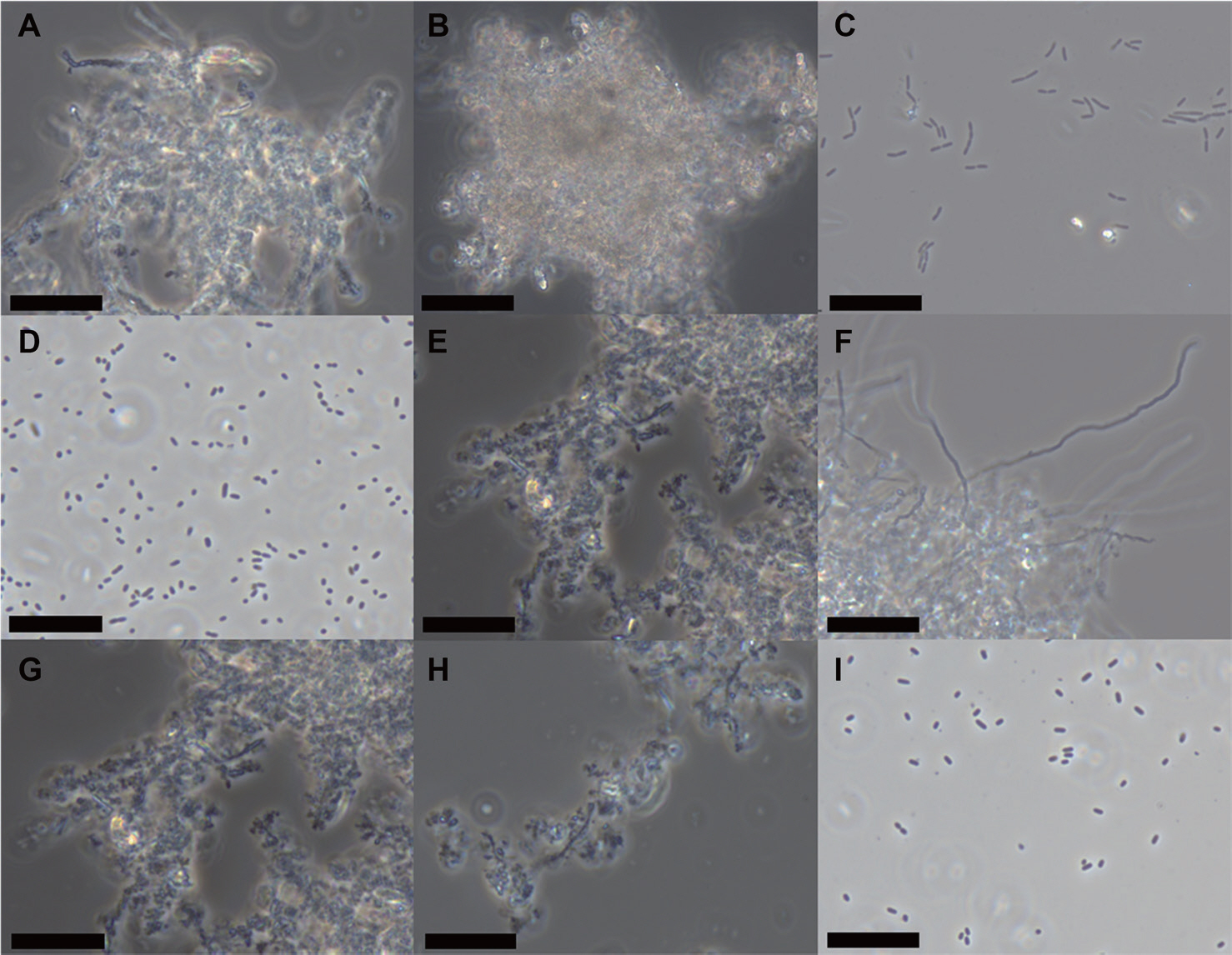
Table 2.
Validation of spore formation ability under microsco-py as affected by nitrogen and carbon sources
| Type | Sources | Sporulation a | |
|---|---|---|---|
| SP6C4 | S8 | ||
| Nitrogen | L-asparagine | - | Y |
| L-cysteine | - | Y | |
| L-glutamic acid | Y | Y | |
| L-proline | Y | Y | |
| L-Valine | - | Y | |
| Peptone | -ah | Y | |
| Carbon | Starch | - | - |
| Yeast extract | - | Y | |
본 연구에서는 기 확보된 유용 방선균(Streptomyces)속 2종을 작물 유래 친환경 미생물 제제로 개발함에 있어 유익성을 제공하고자 그 특성을 구명하고 적합 배양 조건을 확보하려고 하였다. 그 결과 S8과 SP6C4 방선균은 PGPR의 효능이 서로 상이한 특성을 활용하여 혼합 배양 후보군으로서 그 기능이 더 높게 평가될 것으로 생각된다. 탄소원으로 yeast extract 또는 mannitol, 질소원으로는 L-cystein을 기본 영향원으로 사용할 경우, S8과 SP6C4의 공배양에 효율적인 결과를 도출할 것으로 생각된다. 하지만 두 방선균의 생육 최적 산도조건이 S8은 pH 10, SP6C4는 pH 7.2 상이하여 이에 대한 후속 연구 수행이 요구된다. 또한 미생물 제제로서 개발함에 있어 포자의 형태로 존재할 경우 장기보관시 안정성이 높아지고 보관이 용이하므로, 본 연구를 통해 선발된 탄소원·질소원을 통하여 포자 형성을 촉진할 수 있으므로 미생물 제제 개발에 기초적 자료를 마련하였다.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






