

서론
재료 및 방법
병원균 분리 및 동정
배지 종류와 광처리에 따른 포자 생산량
식물 재배
접종원 준비
접종 및 병조사
결과 및 고찰
균주 동정 및 최적 포자 생산 조건
Fig. 1
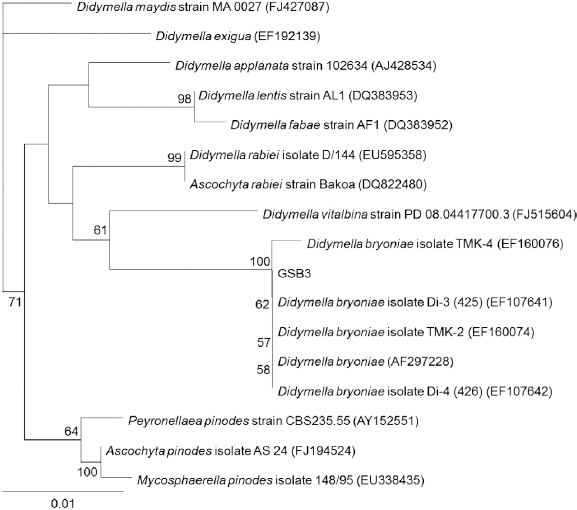
Table 1
| Irradiation condition* | Total incubation period | No. of spores (×107 spores/plate) | ||
|---|---|---|---|---|
|
|
||||
| PDA | V8A | OMA | ||
| A: One-day (12-h light/day) | 8 | 48 bz | 8 ay | 36 bzy |
| A and then two-day darkness | 10 | 182 az | 6 ay | 88 abzy |
| B: Two-day (12-h light/day) | 9 | 152 az | 5 ay | 104 abz |
| B and then two-day darkness | 11 | 153 az | 27 ay | 185 az |
생산 방법에 따른 포자의 병원력 차이
Table 2
Each value represents the mean disease severity±standard deviation of two runs with five replicates each.
Values in the labeled with the same letter in each column are not significantly different in Duncan's multiple range test at P=0.05.
* Seeds of watermelon cultivar were sown and grown in a greenhouse at 25°C±5°C to be growth stage of cotyledone and 1-, 2-, 3-, and 4-fully expanded leaves. Seedlings were inoculated with D. bryoniae by spraying spore suspension of the fungus at a concentration of 5.0×105 spores/ml. The infected plants were incubated in humidity chamber at 25°C for 48 hours and then transferred to a growth chamber at 25°C and 80% relative humidity with 12-hour light a day. Four days after inoculation, disease severity of the plant was measured using percentage of infected leaf area.
수박 22개 품종의 저항성
Fig. 2
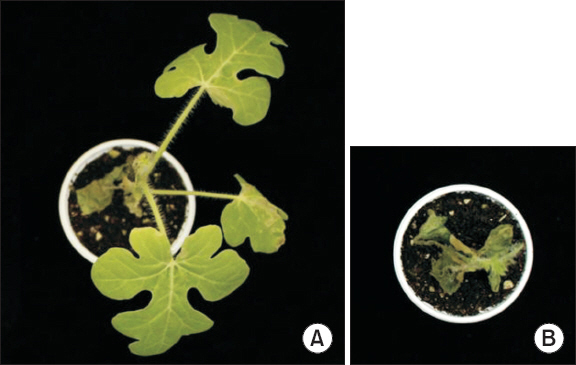
Table 3
Each value represents the mean disease severity±standard deviation. Values in the labeled with the same letter are not significantly different in Duncan's multiple range test at P=0.05.
* Seedlings with 2-fully expanded leaves were inoculated with D. bryoniae by spraying spore suspension of the fungus at a concentration of 5.0×105 spores/ml. The infected plants were incubated in humidity chamber at 25°C for 48 hours and then transferred to a growth chamber at 25°C and 80% relative humidity with 12-hour light a day. Three days after inoculation, disease severity of the plant was measured using percentage of infected leaf area. Experiment was carried out two runs with five replicates each.
습실 처리 기간에 따른 덩굴마름병 발생
Table 4
| Cultivar | Trait | Incubation period in dew chamber | |
|---|---|---|---|
|
|
|||
| 24 h | 48 h | ||
| Wonderfulggul | MR | 8±2.7 az | 19±11 cz |
| Jijonggul | MR | 6±2.2 az | 8±2.7 cz |
| Kokoma | S | 5±0.0 ay | 77±34 abz |
| Seotaja | S | 11±6.5 ay | 84±26 az |
Each value represents the mean disease severity±standard deviation of two runs with five replicates each.
Values in the labeled with the same letter in each column are not significantly different in Duncan's multiple range test at P=0.05. MR, moderately resistant; S, susceptible.
* Seedlings with 2-fully expanded leaves were inoculated with Didymella bryoniae by spraying spore suspension of the fungus at a concentration of 5.0×105 spores/ml. The inoculated plants were incubated in humidity chamber at 25°C for 24 or 48 hours and then transferred to a growth chamber at 25°C and 80% relative humidity with 12-hour light a day. Three days after inoculation, disease severity of the plant was measured using percentage of infected leaf area.
접종 농도에 따른 덩굴마름병 발생
Table 5
Each value represents the mean disease severity±standard deviation of two runs with five replicates each.
Values in the labeled with the same letter in each column are not significantly different in Duncan's multiple range test at P=0.05.
MR, moderately resistant; S, susceptible.
* Seedlings with 2-fully expanded leaves were inoculated with D. bryoniae by spraying spore suspension of the fungus at a concentration of 2.0×104, 1.0×105, 5.0×105, and 2.5×106 spores/ml. The inoculated plants were incubated in humidity chamber at 25°C for 48 hours and then transferred to a growth chamber at 25°C and 80% relative humidity with 12-hour light a day. Three days after inoculation, disease severity of the plant was measured using percentage of infected leaf area.
접종 후 초기 재배온도에 따른 덩굴마름병 발생
Table 6
Each value represents the mean disease severity±standard deviation of two runs with five replicates each.
Values in the labeled with the same letter in each column are not significantly different in Duncan's multiple range test at P=0.05.
MR, moderately resistant; S, susceptible.
* Seedlings with 2-fully expanded leaves were inoculated with D. bryoniae by spraying spore suspension of the fungus at a concentration of 5.0×105 spores/ml. The inoculated plants were incubated in humidity chamber at 20°C, 25°C, and 30°C for 48 hours and then transferred to a growth chamber at 25°C and 80% relative humidity with 12-hour light a day. Three days after inoculation, disease severity of the plant was measured using percentage of infected leaf area.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






