


ņä£ ļĪĀ
ļöĖĻĖ░ņŚÉ ļ░£ņāØĒĢśļŖö ņŗ£ļōżņØīļ│æņØĆ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ ļöĖĻĖ░ļź╝ ņ×¼ļ░░ĒĢśĻ│Ā ņ׳ļŖö ļéśļØ╝ņŚÉņä£ Ļ░Ćņן ļ¼ĖņĀ£Ļ░Ć ļÉśĻ│Ā ņ׳ļŖö ņŻ╝ņÜö ļ│æĒĢ┤ņØ┤ļŗż. ņŗ£ļōżņØīļ│æņØä ņØ╝ņ£╝ĒéżļŖö ļīĆļČĆļČäņØś ļ│æņøÉĻĘĀņØĆ Fusarium oxysporum f. sp. fragariae (Fof)ļĪ£ 1962ļģäņŚÉ ĒśĖņŻ╝ņŚÉņä£ ņĄ£ņ┤ł ļ│┤Ļ│ĀļÉśņŚłņ£╝ļ®░(WinksņÖĆ Williams, 1965), ĻĘĖ Ēøä ņØ╝ļ│Ė(Okamoto ļō▒, 1970), ĒĢ£ĻĄŁ(Kim ļō▒, 1982), ļ®Ģņŗ£ņĮö(Castro-FrancoņÖĆ Davalos-Gonzalez, 1990), ņżæĻĄŁ(Huang ļō▒, 2005), ĒśĖņŻ╝(Golzar ļō▒, 2007), ņŖżĒÄśņØĖ(Ar-royo ļō▒, 2009), ļ»ĖĻĄŁ(Koike ļō▒, 2009, Williamson ļō▒, 2012), ņäĖļź┤ļ╣äņĢä(Stankovi─ć ļō▒, 2014), Ēä░Ēéż(Dinler ļō▒, 2016) ņŚÉņä£ ļ│┤Ļ│ĀļÉśļŖö ļō▒ ņĀä ņäĖĻ│äņĀüņ£╝ļĪ£ ĒÖĢņé░ ņČöņäĖņŚÉ ņ׳ļŗż.
ļöĖĻĖ░ņŚÉ ļ░£ņāØĒĢśļŖö ļ│æņøÉņä▒ņØä Ļ░Ćņ¦ä Fofļź╝ ņŗĀņåŹ ņĀĢĒÖĢĒĢśĻ▓ī ņ¦äļŗ©ĒĢśļŖö ĻĖ░ņłĀņØĆ ļöĖĻĖ░ ņ×¼ļ░░ĒśäņןņŚÉņä£ Ļ░Ćņן ņŗ£ĻĖēĒĢ£ Ļ│╝ņĀ£ņØ┤ļŗż. ņŗØļ¼╝ļ│æ ņ¦äļŗ©ņŚÉ ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ĻĖ░ļ▓ĢņØä ņØ┤ņÜ®ĒĢ£ ļ░®ļ▓ĢņØ┤ Ļ░£ļ░£ļÉśĻ│Ā ņ׳ņ£╝ļ®░(CarboneĻ│╝ Kohn, 1999), ļöĖĻĖ░ ņŗ£ļōżņØīļ│æ ņ¦äļŗ©ņØä ņ£äĒĢ┤ 2013ļģä ņĄ£ņ┤łļĪ£ ņØ╝ļ│ĖņŚÉņä£ Fof ĒŖ╣ņØ┤ņĀü primerĻ░Ć Ļ░£ļ░£ļÉśņŚłļŗż(Suga ļō▒, 2013). Ļ░£ļ░£ļÉ£ primerļŖö ļöĖĻĖ░ ņØ┤ņÖĖņØś ņ×æļ¼╝ņŚÉ ļ░£ņāØĒĢśļŖö formae specialisņØś ļ│æņøÉĻĘĀĻ│╝ ĻĄ¼ļ│äĒĢĀ ņłś ņ׳ņ£╝ļ®░, ņØ╝ļ│ĖņØś ļöĖĻĖ░ņŚÉņä£ ļČäļ”¼ĒĢ£ ļ│æņøÉņä▒Ļ│╝ ļ╣äļ│æņøÉņä▒ Fofļź╝ ĻĄ¼ļ│äĒĢĀ ņłś ņ׳ņ£╝ļéś ļ»ĖĻĄŁ CaliforniaņŚÉņä£ ļČäļ”¼ĒĢ£ FofĻĘĀ ļ¬©ļæÉļź╝ Ļ▓ĆņČ£ĒĢĀ ņłśļŖö ņŚåļŖö ĒĢ£Ļ│äĻ░Ć ņ׳ņŚłļŗż(KoikeņÖĆ Gordon, 2015). Li ļō▒(2014)ļÅä ļöĖĻĖ░ ņ×¼ļ░░ĒżņןņŚÉņä£ ņäĀĒāØļ░░ņ¦ĆņÖĆ real-time PCRņØä ņØ┤ņÜ®ĒĢ┤ ĒåĀņ¢æņØś F. oxysporumņØä Ļ▓ĆņČ£ĒĢĀ ņłś ņ׳ņŚłņ¦Ćļ¦ī FofņØś ĒŖ╣ņØ┤ņĀü ņ¦äļŗ©ņØĆ ļČłĻ░ĆļŖźĒĢ£ ĒĢ£Ļ│äļź╝ ļ│┤ņśĆĻ│Ā ĻĄŁļé┤ņŚÉņä£ļŖö quantitative real-time PCR (qRT-PCR)ņØä ņØ┤ņÜ®ĒĢ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀ ņ¦äļŗ©ĻĖ░ņłĀņØ┤ Ļ░£ļ░£ļÉśĻĖ░ļÅä ĒĢśņśĆļŗż(Hong ļō▒, 2018). ļśÉĒĢ£ Burkhardt ļō▒(2019)ņØĆ ļ»ĖĻĄŁņŚÉ ļ░£ņāØĒĢśļŖö FofņŚÉ ĒŖ╣ņØ┤ņĀüņØĖ Taqman quantitative PCRĻ│╝ isothermal recombinase polymerase amplification assay ĻĖ░ņłĀņØä Ļ░£ļ░£ĒĢśņŚ¼ ņ¦äļŗ©ņŚÉ ņØ┤ņÜ®ĒĢśĻ│Ā ņ׳ļŗż. ĻĘĖļ¤¼ļéś ĻĄŁļé┤ ļ░£ņāØĒĢśļŖö ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØś ņ£ĀņĀäņĀü ĒŖ╣ņä▒ņØ┤ ņĪ░ņé¼ļÉśņ¦Ć ļ¬╗ĒĢ£ ņāüĒā£ņŚÉņä£ļŖö ņĀĢĒÖĢĒ׳ Fof ņ¦äļŗ©ļ░®ļ▓ĢņØä ņĀüņÜ®ĒĢśĻĖ░ ņ¢┤ļĀżņÜ┤ ņĖĪļ®┤ņØ┤ ņ׳Ļ│Ā ņØ┤ņŚÉ ļö░ļźĖ ļ░®ņĀ£ ļ░®ļ▓ĢņØä ņĀüņÜ®ĒĢśĻĖ░ļÅä ņēĮņ¦Ć ņĢŖņØĆ ņŗżņĀĢņØ┤ļŗż.
Ēśäņ×¼ ļöĖĻĖ░ņŚÉ ļō▒ļĪØļÉ£ ņŗ£ļōżņØīļ│æ ņé┤ĻĘĀņĀ£ļŖö dazomatĻ│╝ metam sodiumņØś ĒåĀņ¢æĒøłņ”ØņĀ£ņÖĆ prochloaz manganese, fluxapyroxad+pyraclostrobin, fluxapyroxad+metalaxyl-M, etridiazole+thiophanate-methyl, iprodione+prochloraz manganese, pyribencarb, copper hydroxideņØś 7ņóģņØ┤ ļō▒ļĪØļÉśņ¢┤ ņ׳ļŗż(http://pis.rda.go.kr). Fusarium spp.ņŚÉ ņØśĒĢ£ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņĀ£ļĪ£ļŖö prochloraz (AminiņÖĆ Sidovich, 2010; Shin ļō▒, 2018), prothioconazole (Miller ļō▒, 2020), metconazole (Burlakoti ļō▒, 2010) Ļ│╝ Ļ░ÖņØĆ ņ╣©Ēł¼ņØ┤Ē¢ēņä▒ņØś demethylation inhibitorĻ│ä ņé┤ĻĘĀņĀ£Ļ░Ć ļīĆĒæ£ņĀüņØ┤ļŗż. ļśÉĒĢ£ pyraclostrobin (Cruz Jimenez ļō▒, 2018), thiophanate-methyl (Kim ļō▒, 1982; Masiello ļō▒, 2019; Shin ļō▒, 2018)ļÅä ĒÜ©Ļ│╝Ļ░Ć ņØĖņĀĢļÉśĻ│Ā ņ׳ņ£╝ļéś ņØ┤ļ¤░ ņé┤ĻĘĀņĀ£ļōżņØĆ ņĀĆĒĢŁņä▒ĻĘĀ ļ░£ņāØņØ┤ ļ╣ĀļźĖ ĒŖ╣ņä▒ņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŗż(Petkar ļō▒, 2017). ņĄ£ĻĘ╝ņŚÉļŖö ņŗĀĻĘ£ succinate dehydrogenase inhibitor (SDHI)Ļ│äņØĖ pydiflumetofenņØ┤ F. oxysporum f. sp. niveumņŚÉ ņØśĒĢ£ ņłśļ░Ģ ņŗ£ļōżņØīļ│æņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ļØ╝ ļ│┤Ļ│ĀĒĢśņśĆļŗż(Miller ļō▒, 2020).
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ļŖö ĻĄŁļé┤ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØś ņ£ĀņĀäņĀü ļŗżņ¢æņä▒Ļ│╝ ļ│æņøÉņä▒ņØä ĒÖĢņØĖĒĢśĻ│Ā ļČäļ”¼ĻĘĀņŚÉ ļīĆĒĢ£ ņé┤ĻĘĀņĀ£ ļ░śņØæĻ│╝ ņé┤ĻĘĀņĀ£ņŚÉ ļīĆĒĢ£ ņ×¼ļ░░ĒżņןņŚÉņä£ņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ņĪ░ņé¼ĒĢśņŚ¼ ĒÜ©Ļ│╝ņĀüņØĖ ņŗ£ļōżņØīļ│æ ņ¦äļŗ© ļ░Å ļ░®ņĀ£ņŚÉ ņĀüņÜ®ĒĢśĻĖ░ ņ£äĒĢ┤ ņłśĒ¢ēļÉśņŚłļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ļČäļ”¼ĻĘĀ.
ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀ 27ĻĘĀņŻ╝ļŖö 2003ļģäļČĆĒä░ 2016ļģäļÅÖņĢł ĻĄŁļé┤ ļöĖĻĖ░ ļöĖĻĖ░ņ×¼ļ░░ĒżņןņŚÉņä£ ņŗ£ļōżņØīļ│æņŚÉ ņØ┤ļ│æļÉ£ ņŗØļ¼╝ņ▓┤ļĪ£ļČĆĒä░ ļČäļ”¼ĒĢśņśĆļŗż(Table 1). ļ│æņøÉĻĘĀ ņł£ņłś ļČäļ”¼ļŖö ļöĖĻĖ░ Ļ┤ĆļČĆ, ņŚĮļ│æ, ļ┐īļ”¼ļĪ£ļČĆĒä░ņØś ņØ┤ļ│æņĪ░ņ¦üņØä 70% ņĢīņĮ£ļĪ£ 30ņ┤łĻ░ä Ēæ£ļ®┤ņåīļÅģ Ēøä 3ĒÜī ļ®ĖĻĘĀņłśļĪ£ ņäĖņ▓Ö Ēøä water agar ļ░░ņ¦ĆņŚÉ ņ╣śņāüĒĢśņśĆļŗż. 25┬░C ļ░░ņ¢æĻĖ░ņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æ Ēøä ņ×Éļ×Ć ņäĀļŗ©ņØś ĻĘĀņé¼ļź╝ potato dextrose agar (PDA) ļ░░ņ¦ĆņŚÉņä£ 27┬░C ļ░░ņ¢æ Ēøä ņ×Éļ×Ć colonyņÖĆ ļČäņāØĒżņ×É ĒśĢĒā£ ļ░Å single palide ĒśĢĒā£ļź╝ Ēśäļ»ĖĻ▓Įņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņŚ¼ FofļĪ£ ļÅÖņĀĢĒĢśņśĆļŗż. ļśÉĒĢ£ ļŹö ņ×ÉņäĖĒĢ£ ļÅÖņĀĢņØä ņ£äĒĢ┤ Suga ļō▒(2013)ņØś Fof ĒŖ╣ņØ┤ņĀü primers (FofraF/FofraR primers)ņÖĆ Burkhardt ļō▒(2019)ņØś qRT-PCR probe (Frag_F/Fraf_R/Frag_TaqMan_probe)ļź╝ ņØ┤ņÜ®ĒĢ┤ DNA ņ”ØĒÅŁ ņŚ¼ļČĆļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. qRT-PCRņØĆ CFX96 real-time PCR detection system (Bio-Rad, Hercules, CA, USA)ļź╝ ņØ┤ņÜ®ĒĢ┤ ļČäņäØĒĢśņśĆļŗż. XENO Mastermix (Genotype, Daejeon, Korea)ņŚÉ DNA 1 ╬╝lņÖĆ primersņÖĆ probeļź╝ Ļ░üĻ░ü 1 ╬╝lņö® ļäŻĻ│Ā, 16 ╬╝lņØś ļ®ĖĻĘĀņłśļź╝ ļäŻņ¢┤ ņĄ£ņóģļ░śņØæņĢĪņØä 20 ╬╝lļĪ£ ņĪ░ņä▒ĒĢśņśĆļŗż. PCR ņĪ░Ļ▒┤ņØĆ 95┬░CņŚÉņä£ pre-denaturation 3ļČä, 95┬░CņŚÉņä£ denaturation 15ņ┤ł, 62┬░CņŚÉņä£ annealing 30ņ┤ł, 72┬░CņŚÉņä£ extension 30ņ┤łņØś ņĪ░Ļ▒┤ņ£╝ļĪ£ 50 cyclesņØś PCRņØä ņłśĒ¢ēĒĢśņśĆļŗż.
ļ│æņøÉņä▒ Ļ▓ĆņĀĢ.
ļöĖĻĖ░ņŚÉņä£ ļČäļ”¼ĒĢ£ ņŗ£ļōżņØīļ│æĻĘĀņŚÉ ļīĆĒĢ£ ļ│æņøÉņä▒ Ļ▓ĆņĀĢņØĆ ļöĖĻĖ░ņŚ░ĻĄ¼ņåīņØś ļ│æ Ļ▓ĆņĀĢ ļ╣äļŗÉĒĢśņÜ░ņŖżņŚÉņä£ ņŗżņŗ£ĒĢśņśĆļŗż. ņŗ£ĒŚś ĒÆłņóģņØĆ ņĪ░ņ¦üļ░░ņ¢æļ¼śļĪ£ļČĆĒä░ ņ£Īļ¼śĒĢ£ ņäżĒ¢ź ĒÆłņóģņØä ņŗ£ĒŚśņŚÉ ņØ┤ņÜ®ĒĢśņśĆņ£╝ļ®░, ņāüĒåĀļŖö ļöĖĻĖ░ņĀäņÜ®ņāüĒåĀ(ņĮöņĮöĒö╝ĒŖĖ:Ēö╝ĒŖĖļ¬©ņŖż:ĒÄäļØ╝ņØ┤ĒŖĖ=65:17:10, Purumi, SeoulBio, Eumseong, Korea)ļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. ņŗ£ļōżņØīļ│æĻĘĀņØĆ minimal medium broth ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 27┬░C, 120 rpmņØś shaking incubatorņŚÉņä£ 7ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ļ░░ņ¢æņŚ¼ņĢĪņØĆ ļ®ĖĻĘĀļÉ£ 4Ļ▓╣ņØś Ļ▒░ņ”łļĪ£ ĻĘĀņé¼ļź╝ Ļ▒░ļźĖ Ēøä hemocytometerļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäņāØĒżņ×ÉĻ░Ć 1├Ś105 conidia/ml ļåŹļÅäļĪ£ ņĪ░ņĀĢĒĢśņŚ¼ ļ│æņøÉņä▒ Ļ▓ĆņĀĢņŚÉ ņØ┤ņÜ®ĒĢśņśĆļŗż. ņŗØļ¼╝ņ▓┤ņØś ļ┐īļ”¼ļŖö 5 cmļ¦ī ļé©ĻĖ░Ļ│Ā Ļ░Ćņ£äļĪ£ ņ×ÉļźĖ ļŗżņØī ņŗ£ļōżņØīļ│æĻĘĀ ļ░░ņ¢æņĢĪņŚÉ 10ļČäĻ░ä ļ┐īļ”¼ ņ╣©ņ¦Ć Ēøä ņāüĒåĀĻ░Ć ļŗ┤ĻĖ┤ ĒżĒŖĖ(ņ¦üĻ▓Į 10 cm)ņŚÉ ņØ┤ņŗØĒĢśņśĆļŗż. ļ╣äļŻī ņŗ£ļ╣äļŖö ņØ┤ņŗØ 1ņŻ╝ņØ╝ ĒøäļČĆĒä░ Ferticare (20-20-20, Yara, Oslo, Norway) 1,000ļ░░ņĢĪņØä 1ņŻ╝ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ ņŻ╝ļŗ╣ 50 mlņö® Ļ┤ĆņŻ╝ĒĢśņśĆļŗż. ņŗØļ¼╝ņ▓┤ļŖö Ļ░üĻ░üņØś ņŗ£ļōżņØīļ│æĻĘĀļŗ╣ 5ņŻ╝ļź╝ ņŗ£ĒŚśņŚÉ ņØ┤ņÜ®ĒĢśņśĆļŗż. ņØ┤ļ│æņ¦ĆņłśļŖö ņŗ£ļōżņØīļ│æĻĘĀ ņĀæņóģ 52ņØ╝ņŚÉ ņĪ░ņé¼ĒĢśņśĆļŗż. ņØ┤ļ│æņ¦ĆņłśļŖö 0 ļ¼┤ļ│æņ¦Ģ, 1 ņŗĀņŚĮĒÖ®ņāē, ĒĢśņŚĮņŚĮļ│æ ņĀüņāē, 2 ņ¦Øņ×Ä, 3 1/2ņŗ£ļō”, 4 ņĀäņ▓┤ņŗ£ļō”, 5 Ļ│Āņé¼ņØ┤ļŗż(Nam ļō▒, 2005).
ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ļČäņäØ.
ļöĖĻĖ░ņŚÉņä£ ļČäļ”¼ĒĢ£ 27ĻĘĀņŻ╝ņŚÉ ļīĆĒĢ£ DNA sequencing ļČäņäØņØä ņŗżņŗ£ĒĢśņśĆņ£╝ļ®░ ņŗ£ļōżņØīļ│æĻĘĀņØś DNA ļČäļ”¼ļŖö Park ļō▒(2005)ņØś ļ░®ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. Nuclear ribosomal inter-genic spacer region (IGS)ņÖĆ translation elongation factor EF-l ╬▒ gene (tef1) genesņØĆ species ņłśņżĆņŚÉņä£ Ļ░üĻ░üņØś ĻĘĀņŻ╝ļź╝ ļČäņäØĒĢśĻĖ░ ņ£äĒĢ┤ ņ”ØĒÅŁĒĢśņśĆļŗż. IGSņÖĆ tef1 genesņØś PCR ņ”ØĒÅŁņØĆ iNL11/CNSa/iCNS11/NLa (OŌĆÖDonnell ļō▒, 2009)Ļ│╝ EF1-728F/EF1-986R (CarboneĻ│╝ Kohn, 1999)ļź╝ ņØ┤ņÜ®ĒĢśņśĆļŗż. PCR ļ░śņØæņØĆ ņØ┤ņĀäņŚÉ ļ│┤Ļ│ĀļÉ£ ļ░®ļ▓ĢņŚÉ ņØśĒĢ┤ CFX96 thermal cycler (Bio-Rad)ļĪ£ ņŗżņŗ£ĒĢśņśĆļŗż. PCR ņé░ļ¼╝ņØĆ PCRquick-spin PCR Product Purification Kit (iNtRON BioTechnology, Seongnam, Korea)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļČäļ”¼ Ēøä Bioneer Corporation (Cheongwon, Korea)ņŚÉ sequencing ļČäņäØņØä ņØśļó░ĒĢśņśĆļŗż. SequencesļŖö MEGA 5 software (Tamura ļō▒, 2011)ļź╝ ņØ┤ņÜ®ĒĢ┤ ĒÄĖņ¦æ Ēøä MAFFT v7 (KatohņÖĆ Standley, 2013)ļĪ£ alignĒĢśņśĆļŗż. Combined data set (IGS+tef1)ļź╝ ņØ┤ņÜ®ĒĢ£ maxi-mum likelihood ļČäņäØņØĆ GTR├ŠG modelĻ│╝ 1,000 bootstrap replicatesņØä ņØ┤ņÜ®ĒĢ┤ RAxML version 7.03 (Bioinformatics Insti-tute, Singapore) (Stamatakis, 2006)ļĪ£ ļČäņäØĒĢśņśĆļŗż.
ņé┤ĻĘĀņĀ£ ļ░śņØæ.
ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņŚÉ ļīĆĒĢ£ ņé┤ĻĘĀņĀ£ ļ░śņØæ ņĪ░ņé¼ņŚÉ ņé¼ņÜ®ļÉ£ ņé┤ĻĘĀņĀ£ļŖö prochloraz (a.i. 25%, EC), metconazole (a.i. 20%, SC), pyraclostrobin (a.i. 11%, SC), fluxapyroxad (a.i. 15.3%, SC), dimethomorph (a.i. 25%, WP), thiophanate-methyl (a.i. 70%, WP), copper hydroxide (a.i. 77%, WP)ņØś 7ņóģņØ┤ņŚłļŗż. Ļ░ü ņé┤ĻĘĀņĀ£ļŖö PDA ļ░░ņ¦ĆņŚÉņä£ ņĄ£ņóģļåŹļÅä(a.i. ĻĖ░ņżĆ)Ļ░Ć 0.001ņŚÉņä£ 100 ╬╝g/mlĻ╣īņ¦Ć ņé┤ĻĘĀņĀ£ļ│ä ļåŹļÅäļź╝ ļŗ¼ļ”¼ĒĢśņŚ¼ ņ▓©Ļ░ĆĒĢśņśĆļŗż. ņŗżĒŚśņŚÉ ņé¼ņÜ®ĒĢ£ ņé┤ĻĘĀņĀ£ļź╝ ļ®ĖĻĘĀņ”ØļźśņłśņŚÉ ĒśäĒāüĒĢ£ Ēøä, PDA ļ░░ņ¦ĆņŚÉ ņĀĢĒĢ┤ņ¦ä ļåŹļÅäĻ░Ć ļÉśļÅäļĪØ ļ¦×ņČöņ¢┤ ņ▓©Ļ░ĆĒĢśņśĆņ£╝ļ®░ ņäĖĻĘĀ ņśżņŚ╝ ļ░®ņ¦Ćļź╝ ņ£äĒĢ┤ streptomycin 300 ╬╝g/mlļź╝ ļ░░ņ¦ĆņŚÉ ņ▓©Ļ░ĆĒĢśņśĆļŗż. ņé┤ĻĘĀņĀ£Ļ░Ć ļōżņØĆ ļ░░ņ¦ĆņŚÉ ņŗ£ļōżņØīļ│æĻĘĀņØä ņĀæņóģņØä ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņżĆļ╣äĒĢ£ FofņØś ĻĘĀņé¼ņäĀļŗ©ņŚÉņä£ ĻĘĀņé¼ ņĪ░Ļ░ü(Ļ░ĆļĪ£├ŚņäĖļĪ£, 1├Ś1 mm)ņØä ļ¢╝ņ¢┤ ļé┤ņ¢┤ ņé┤ĻĘĀņĀ£ļź╝ ņ▓©Ļ░ĆĒĢ£ PDA ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśĻ│Ā, 27┬░CņØś ņĢöņāüĒā£ņŚÉņä£ 5ņØ╝Ļ░äļ░░ņ¢æĒĢśņśĆņ£╝ļ®░ ņ▓śļ”¼ļÉ£ ļ¬©ļōĀ ļåŹļÅäņŚÉņä£ 3ļ░śļ│Ąņ£╝ļĪ£ ņ¦äĒ¢ēļÉśņŚłļŗż. ņé┤ĻĘĀņĀ£ ĒÜ©Ļ│╝ļŖö 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņé┤ĻĘĀņĀ£ ļ░░ņ¦ĆņŚÉņä£ ņ×Éļ×Ć ļ│æņøÉĻĘĀņØśĻĘĀņ┤ØņØś ņ¦üĻ▓ĮĻ│╝ ļ¼┤ņ▓śļ”¼ ļ░░ņ¦ĆņŚÉņä£ ļ│æņøÉĻĘĀ ĻĘĀņ┤ØņØś ņ¦üĻ▓ĮņØä ņĪ░ņé¼ĒĢśĻ│Ā, ņĢäļל ņŗØņŚÉ ņØśĒĢ┤ņä£ ĻĘĀņé¼ņāØņן ņ¢ĄņĀ£ĒÜ©Ļ│╝ļź╝ ĻĄ¼ĒĢśņŚ¼ ļ╣äĻĄÉĒĢśņśĆļŗż.
ņé┤ĻĘĀņĀ£ņØś ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ĒÜ©Ļ│╝.
ļöĖĻĖ░ ņ£Īļ¼śĒżņןņŚÉņä£ ņé┤ĻĘĀņĀ£ļ│ä ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. ņé┤ĻĘĀņĀ£ļŖö boscalid+fludioxonil (a.i. 23.5+5%, S1 SC, FarmHannong, Seoul, Korea) 1,000ļ░░, fluxapyroxad+pyraclostrobin (a.i. 4.6+8.3%, Mirivon SC, FarmHannong) 2,000ļ░░, prochloraz manganese (a.i. 50%, Sporgon WP, HanKookSamGong, Seoul, Korea) 2,000ļ░░, copper hydroxide (a.i. 40%, Daeyucucide SC, Daeyu, Seoul, Korea) 500ļ░░ļĪ£ ņŻ╝ļŗ╣ 100 mlņö® 1ņŻ╝ņØ╝ Ļ░äĻ▓® 3ĒÜī ĻĘ╝ļČĆ Ļ┤ĆņŻ╝ ņ▓śļ”¼ĒĢśņśĆļŗż. ņŗØļ¼╝ņ▓┤ļŖö ņäżĒ¢źĒÆłņóģņ£╝ļĪ£ ņĪ░ņ¦üļ░░ņ¢æļ¼śļĪ£ļČĆĒä░ ņ£Īļ¼śĒĢ£ ņ×Éļ¼śļź╝ ņé¼Ļ░üĒżĒŖĖ(55├Ś19.6├Ś14.5 cm, 10 l)ļŗ╣ 5ņŻ╝ņö® ļöĖĻĖ░ņĀäņÜ®ņāüĒåĀņŚÉ ņĀĢņŗØĒĢśņśĆĻ│Ā ņ▓śļ”¼ļŗ╣ 4ļ░śļ│Ąņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż. 1ņ░©ņŗ£ĒŚśņŚÉņä£ ņŗ£ļōżņØīļ│æĻĘĀ ņĀæņóģņØĆ ņ£äņØś ļ│æņøÉņä▒ Ļ▓ĆņĀĢ ļ░®ļ▓ĢņŚÉ ļÅÖņØ╝ĒĢśĻ▓ī ņĀæņóģņøÉņØä ļ¦īļōżņ¢┤ Ēżņ×ÉĒśäĒāüņĢĪ(1├Ś106 conidia/ml)ņØä ņŻ╝ļŗ╣10 mlņö® 4ņøö 25ņØ╝ņŚÉ ĻĘ╝ļČĆņĀæņóģĒĢ£ Ēøä ņé┤ĻĘĀņĀ£ļŖö 4ņøö 28ņØ╝, 5ņøö 4ņØ╝, 5ņøö 12ņØ╝ 3ĒÜī ņ▓śļ”¼ĒĢśņśĆļŗż. 2ņ░© ņŗ£ĒŚśņŚÉņä£ļŖö ņĀäļģäļÅä ņŗ£ļōżņØīļ│æņØ┤ļ░£ņāØļÉ£ ņØ┤ļ│æņāüĒåĀņŚÉ ņäżĒ¢źĒÆłņóģņØä 4ņøö 27ņØ╝ņŚÉ ņĀĢņŗØ Ēøä 6ņøö 20ņØ╝, 6ņøö 27ņØ╝, 7ņøö 4ņØ╝ņŚÉ ņé┤ĻĘĀņĀ£ļź╝ ņ▓śļ”¼ĒĢśņśĆļŗż. ņØ┤ļ│æņ¦ĆņłśļŖö ņ£äņÖĆĻ░ÖņØ┤ Nam ļō▒(2005)ņØś ļ░®ļ▓Ģņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆĻ│Ā ļ░®ņĀ£ĒÜ©Ļ│╝ļŖö ļŗżņØīĻ│╝ Ļ░ÖņØĆ ņŗØņŚÉ ļö░ļØ╝ Ļ│äņé░ĒĢśņśĆļŗż.
ņ▓śļ”¼Ļ░ä ļ╣äĻĄÉļŖö CoStatĒåĄĻ│ä ĒöäļĪ£ĻĘĖļש(CoHort software, Berkeley, CA, USA)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ DuncanŌĆÖs multiple range test (P<0.05)Ļ▓ĆņĀĢņØä ņŗżņŗ£ĒĢśņśĆļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ļČäļ”¼ĻĘĀ ĒŖ╣ņ¦Ģ.
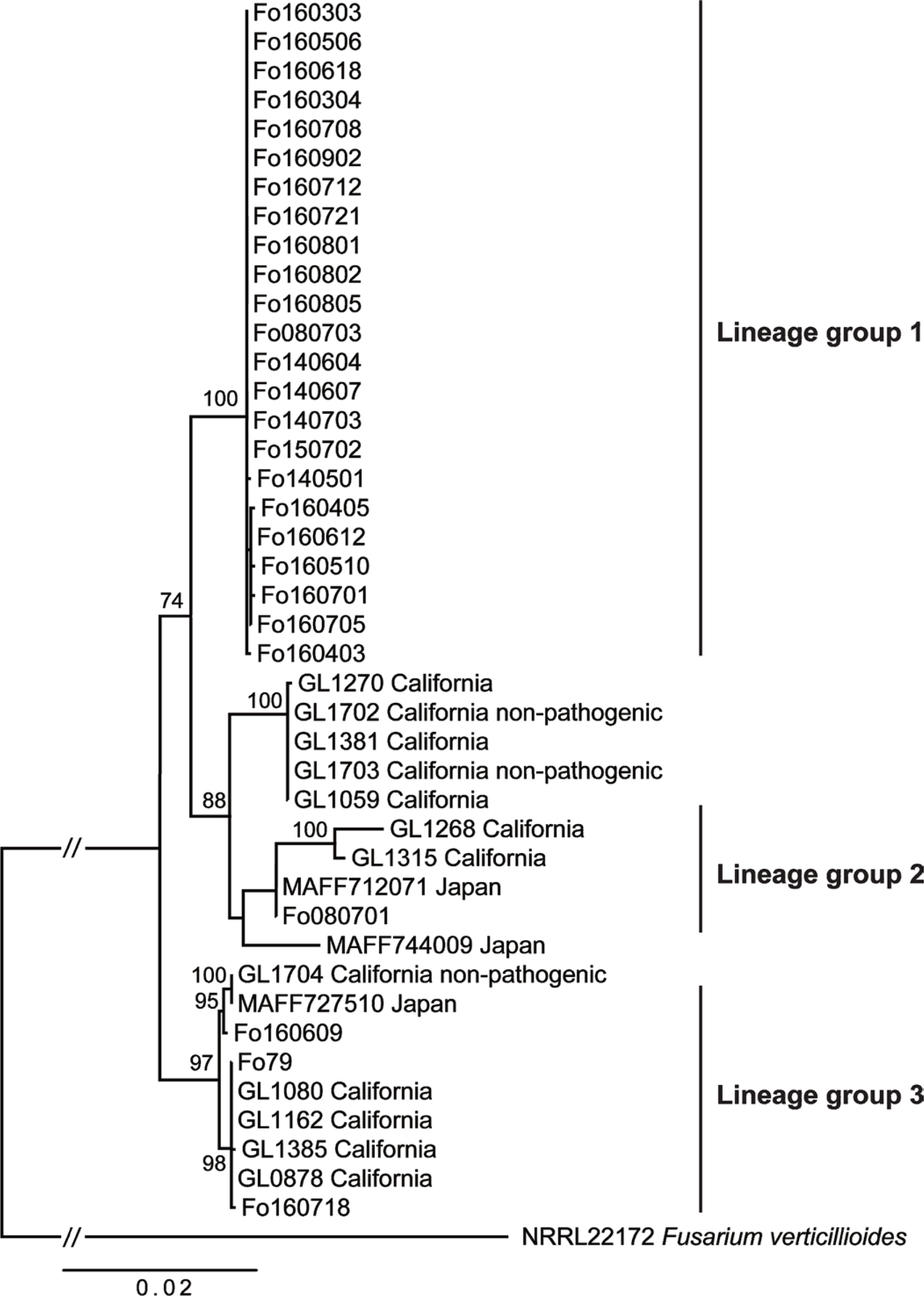
ļöĖĻĖ░ņŚÉņä£ ļČäļ”¼ĒĢ£ ņŗ£ļōżņØīļ│æĻĘĀņØś ĒŖ╣ņ¦ĢņØĆ Table 1Ļ│╝ Ļ░Öļŗż. ļČäļ”¼ĻĘĀņØĆ 2003ļģäļČĆĒä░ 2016ļģäĻ╣īņ¦Ć ĻĄŁļé┤ ļöĖĻĖ░ ņ×¼ļ░░ĒżņןņŚÉņä£ ņŗ£ļōżņØī ņ”ØņāüņØä ļ│┤ņØ┤ļŖö ļöĖĻĖ░ņØś Ļ┤ĆļČĆ, ņŚĮļ│æ, ļ┐īļ”¼ņŚÉņä£ ļČäļ”¼ļÉśņŚłļŗż. ļČäļ”¼ĻĘĀņØś colony ņāēņØĆ PDA ļ░░ņ¦ĆņŚÉņä£ ņŚ░ĒĢ£ ļČäĒÖŹņāēņØś ĻĘĀņé¼ļź╝ ĒśĢņä▒ĒĢśņśĆĻ│Ā ņ┤łņāØļŗ¼ļ¬©ņ¢æņØś ņåīĒśĢ ļ░Å ļīĆĒśĢ ļČäņāØĒżņ×ÉņÖĆ single palideĒśĢņä▒ņ£╝ļĪ£ ĒśĢĒā£ņĀü ļÅÖņĀĢņØä ĒĢśņśĆļŗż(Nelson ļō▒, 1983). ļČäļ”¼ĻĘĀņØĆ Suga ļō▒(2013)ņØ┤ Ļ░£ļ░£ĒĢ£ Fof ĒŖ╣ņØ┤ņĀü primerļĪ£ DNA ņ”ØĒÅŁĒĢ£ Ļ▓░Ļ│╝ Fo080701ĻĘĀņŻ╝(lineage 2ĻĘĖļŻ╣)ļź╝ ņĀ£ņÖĖĒĢ£ ļ¬©ļōĀ ļČäļ”¼ĻĘĀņŚÉņä£ ņ”ØĒÅŁņØ┤ ĒÖĢņØĖļÉśņŚłļŗż(data not shown). ļ░śļ®┤ Burkhardt ļō▒(2019)ņØ┤ Ļ░£ļ░£ĒĢ£ qRT-PCR probeļŖö Fo080701, Fo160609, Fo79, Fo1607818 (lineage 2, 3ĻĘĖļŻ╣)ņØś 4ĻĘĀņŻ╝ļ¦ī Ļ▓ĆņČ£ļÉśņŚłļŗż. ļČäļ”¼ĻĘĀņØś ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ņØä ņ£äĒĢ£ IGSņÖĆ EF-1╬▒ sequencesņØś ņ£ĀņŚ░Ļ┤ĆĻ│ä ļČäņäØĻ▓░Ļ│╝ ĻĄŁļé┤ ņŗ£ļōżņØīļ│æĻĘĀņØĆ 3Ļ░Ćņ¦Ć ĻĘĖļŻ╣ņØś lineagesļź╝ ļ│┤ņśĆļŗż(Fig. 1). ļīĆļČĆļČäņØś ļČäļ”¼ĻĘĀņØĆ DNA lineage 1Ļ│╝ 3ņŚÉ ņåŹĒĢśņśĆņ£╝ļ®░ ļŗ©ņ¦Ć Fo080701ņØś 1ĻĘĀņŻ╝ļ¦ī lineage 2ņŚÉ ņåŹĒ¢łļŗż. 3ĻĘĀņŻ╝ Fo160801, Fo080701, Fo79ņØś IGSņÖĆ EF-1╬▒ sequencesļŖö GenBankņŚÉ ļō▒ļĪØĒĢśņśĆļŗż(MT671188, MT671185; MT671189, MT671186; MT671190, MT671187). ĻĄŁļé┤ DNA lineage 1 ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö ĻĘĀņŻ╝ļŖö ļ»ĖĻĄŁņØ┤ļéś ņØ╝ļ│ĖņØś ņŗ£ļōżņØīļ│æĻĘĀĻ│╝ ļŗżļźĖ ĻĘĖļŻ╣ņØä ĒśĢņä▒ĒĢśņśĆļŗż. ĻĄŁļé┤ DNA lineage 3ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö ĻĘĀņŻ╝ļŖö ļ»ĖĻĄŁ CaliforniaņØś ļČäļ”¼ĻĘĀ somatic compatibility group (SCG) 1ĻĘĖļŻ╣ ļ░Å ņØ╝ļ│ĖņØś ņŗ£ļōżņØīļ│æĻĘĀ MAFF727510Ļ│╝ Ļ░ÖņØĆ ĻĘĖļŻ╣ņØä ĒśĢņä▒ĒĢśņśĆļŗż. ļ░śļ®┤ ĻĄŁļé┤ DNA lineage 2ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö Fo080701ņØĆ ņØ╝ļ│ĖņØś ņŗ£ļōżņØīļ│æĻĘĀ MAFf712071Ļ│╝ Ļ░ÖņØĆ ĻĘĖļŻ╣ņØä ļ│┤ņśĆļŗż. ļČäļ”¼ĻĘĀņØś ļöĖĻĖ░ ņäżĒ¢ź ĒÆłņóģņŚÉ ļīĆĒĢ£ ļ│æņøÉņä▒ Ļ▓ĆņĀĢ Ļ▓░Ļ│╝ ļČäļ”¼ĻĘĀņŚÉ ļö░ļØ╝ ņĀĢļÅäņØś ņ░©ņØ┤ļŖö ņ׳ņ¦Ćļ¦ī ļ¬©ļōĀ ļČäļ”¼ĻĘĀņØĆ ņäżĒ¢ź ĒÆłņóģņŚÉ ļ│æņøÉņä▒ņØ┤ ĒÖĢņØĖļÉśņŚłļŗż. ĒŖ╣Ē׳, Fo160506Ļ│╝ Fo160902ļŖö ņØ┤ļ│æņ¦ĆņłśĻ░Ć 4 ņØ┤ņāüņØś ļåÆņØĆ ļ│æņøÉņä▒ņØä ļ│┤ņśĆņ£╝ļ®░ lineage 1Ļ│╝ 3ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö ļČäļ”¼ĻĘĀņØĆ ļīĆļČĆļČä ļåÆņØĆ ņØ┤ļ│æņ¦Ćņłśļź╝ ļ│┤ņØĖ ļ░śļ®┤ 2ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö ĻĘĀņŻ╝ļŖö ļé«ņØĆ ņØ┤ļ│æņĀĢļÅäļź╝ ļ│┤ņśĆļŗż.
Suga ļō▒(2013)ņØś ņŗ£ļōżņØīļ│æ ĒŖ╣ņØ┤ņĀü primerļŖö Fo080701ņØä ņĀ£ņÖĖĒĢ£ ĻĄŁļé┤ ļīĆļČĆļČäņØś ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░ ļ»ĖĻĄŁ ĻĘĀņŻ╝ņŚÉņä£ļŖö SCG1ņØś ĒĢ£ ĻĘĀņŻ╝ņÖĆ SCG2ņŚÉ ņåŹĒĢśļŖö ĻĘĀņŻ╝Ļ░Ć ņ”ØĒÅŁļÉśņ¦Ć ņĢŖļŖö Ļ▓░Ļ│╝ļź╝ ļ│┤ņśĆļŗż(Henry ļō▒, 2017). ļśÉĒĢ£ Pastrana ļō▒(2017)ļÅä FPOST-140, G2-4 (Spain), ATCC 18138 (Australia), NRRL 26438 (South Carolina) ĻĘĀņŻ╝ļÅä ņ”ØĒÅŁļÉśņ¦Ć ņĢŖļŖö Ļ▓░Ļ│╝ļź╝ ļ│┤ņŚ¼ Fof ĒŖ╣ņØ┤ņĀü primerļŖö Fof ļ¬©ļæÉļź╝ Ļ▓ĆņČ£ĒĢśņ¦ĆļŖö ļ¬╗ĒĢśļŖö ĒĢ£Ļ│äļź╝ ļ│┤ņśĆņ£╝ļ®░ FofļŖö ļéśļØ╝ļ│ä ņŗ£ļōżņØīļ│æĻĘĀņØś ļŗżņ¢æĒĢ£ populationņØ┤ ņĪ┤ņ×¼ĒĢ£ļŗżļŖö Ļ▓āņØä ņ£ĀņČöĒĢĀ ņłś ņ׳ņŚłļŗż. ļ░śļ®┤ Fof ĒŖ╣ņØ┤ņĀü primerļĪ£ Ļ▓ĆņČ£ĒĢĀ ņłś ņŚåļŖö Fofļź╝ Ļ▓ĆņČ£ĒĢśĻĖ░ ņ£äĒĢ┤ Ļ░£ļ░£ļÉ£ probe (Burkhardt ļō▒, 2019)ļŖö ĻĄŁļé┤ lineage 2ņÖĆ 3ļ¦ī Ļ▓ĆņČ£ņØ┤ Ļ░ĆļŖźĒĢ£ ĒĢ£Ļ│äļź╝ ļ│┤ņśĆļŗż. ļö░ļØ╝ņä£ ĻĄŁļé┤ ļ░£ņāØĒĢśļŖö FofļŖö Burkhardt ļō▒(2019)ņØś probeļ│┤ļŗżļŖö Fof ĒŖ╣ņØ┤ņĀü primerļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ļ▓ĆņČ£ĒĢśļŖö Ļ▓āņØ┤ ĒÜ©Ļ│╝ņĀüņØ┤ļ®░ Ļ░£ļ░£ļÉ£ primerļź╝ ņØ┤ņÜ®ĒĢ£ real-time PCRņÜ® probe Ļ░£ļ░£ļÅä ĒĢäņÜöĒĢśļŗż.
ļöĖĻĖ░ ņŗ£ļōżņØīļ│æņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ļ│æņøÉĻĘĀņØĆ ļīĆļČĆļČä FofņØ┤ļ®░ F. oxysporumņØś formae specialisļŖö ņĀ£ĒĢ£ļÉ£ ĻĖ░ņŻ╝ ļ▓öņ£äļź╝ Ļ░Ćņ¦Ćļ®░ ņØ┤ ļ│æņøÉĻĘĀņØĆ ļŗ©ņ¦Ć ļöĖĻĖ░ņŚÉļ¦ī ļ░£ņāØĒĢśļŖö ĒŖ╣ņ¦ĢņØ┤ ņ׳ļŗż(Kodama, 1974). ņĄ£ĻĘ╝ ļČäņ×ÉņāØļ¼╝ĒĢÖņĀü ĻĖ░ļ▓ĢņØä ņØ┤ņÜ®ĒĢ£ ņŗØļ¼╝ļ│æņøÉĻĘĀ ļČäļźśĻ░Ć Ļ░£ļ░£ļÉśņŚłņ£╝ļ®░ ļ│æņøÉĻĘĀ DNAņØś IGSņÖĆ elongation factorņśüņŚŁņØä sequencingĒĢśņŚ¼ ņĀĢĒÖĢĒ׳ F. oxysporumņØś formae specialisĻ╣īņ¦Ć ĻĄ¼ļČäĒĢĀ ņłś ņ׳Ļ▓ī ļÉśņŚłļŗż(OŌĆÖDonnell ļō▒, 2009). ĻĄŁļé┤ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØś IGSņÖĆ EF-1╬▒ sequenceļČäņäØņ£╝ļĪ£ FofļŖö ļīĆļČĆļČä lineage 1Ļ│╝ 3ņŚÉ ņåŹĒĢśļ®░ ņØ┤ļōż ĻĘĖļŻ╣ņØĆ ņäżĒ¢ź ĒÆłņóģņŚÉ ļīĆĒĢ£ ļ│æņøÉņä▒ļÅä ļåÆņØĆ Ļ▓ĮĒ¢źņØä ļ│┤ņØ┤ļŖö ĒŖ╣ņ¦ĢņØä ļ│┤ņśĆļŗż. Kim ļō▒(2017)ņŚÉ ņØśĒĢśļ®┤ ĻĄŁļé┤ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØĆ PCR-restriction fragment length ploymorphism ļČäņäØĻ▓░Ļ│╝ 5ĻĘĖļŻ╣ņ£╝ļĪ£ ņäĖļČäļÉśņŚłĻ│Ā ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ņØĆ ņ×¼ļ░░ ņ¦ĆņŚŁĻ│╝ Ļ┤ĆļĀ©ļÉ£ļŗżĻ│Ā ĒĢśņśĆļŗż. Kim ļō▒(2017)ņØ┤ ļ░£Ēæ£ĒĢ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØĆ IGSņÖĆ EF-1╬▒ sequence ļČäņäØĻ▓░Ļ│╝ ļ¬©ļæÉ lineage 1ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö Ļ▓░Ļ│╝ļź╝ ļ│┤ņŚ¼ ĻĄŁļé┤ņŚÉļŖö ņØ┤ ĻĘĖļŻ╣ņØ┤ ņÜ░ņĀÉņĀüņ£╝ļĪ£ ļČäĒżĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ│┤ņØĖļŗż(data not shown). ļ»ĖĻĄŁņØś ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņØĆ vegetative compatibility group ļČäņäØ Ļ▓░Ļ│╝ 3Ļ░£ņØś ĻĘĖļŻ╣ņ£╝ļĪ£ ņäĖļČäļÉśņŚłņ£╝ļ®░(Henry ļō▒, 2017), ĒśĖņŻ╝ņØś ņŗ£ļōżņØīļ│æĻĘĀņØĆ EF-1╬▒ sequence ļČäņäØ ņŗ£ ļ¦ÄņØĆ lineageĻĘĖļŻ╣ņØä ĒśĢņä▒ĒĢśņśĆļŗż(Paynter ļō▒, 2016). ņĄ£ĻĘ╝, ļ»ĖĻĄŁņÖĆ ĒśĖņŻ╝ņŚÉņä£ļŖö FofņØś population dynamicsņŚÉ ļīĆĒĢ£ ņŚ░ĻĄ¼ļź╝ ņłśĒ¢ēĒĢśņŚ¼ ņŗ£ļōżņØīļ│æ ļ░£ņāØĻĄŁ Ļ░ä FofņØś ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļ░ØĒ׳ļĀżļŖö ņŚ░ĻĄ¼Ļ░Ć ĒÖ£ļ░£Ē׳ ņłśĒ¢ēļÉśĻ│Ā ņ׳ļŗż(Henry ļō▒, 2017; Paynter ļō▒, 2016). ņ£äņØś Ļ▓░Ļ│╝ļĪ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æ ļ░£ņāØņØ┤ ļåÆņØĆ ļ»ĖĻĄŁ, ĒśĖņŻ╝, ņØ╝ļ│Ė, ņŖżĒÄśņØĖ ļō▒ņØś ņŗ£ļōżņØīļ│æĻĘĀņØĆ ļÅģĒŖ╣ĒĢ£ ņ£ĀņĀäņĀü ļŗżņ¢æņä▒ņØä ĒśĢņä▒ĒĢśĻ│Ā ņ׳ņ£╝ļ®░ ņČöĒøä ņØ┤ļōż ņé¼ņØ┤ņØś ņ£ĀņŚ░Ļ┤ĆĻ│äļź╝ ļ░ØĒ׳ļŖö Ļ▓āņØ┤ ņŚ░ĻĄ¼ļÉśņ¢┤ņĢ╝ ĒĢĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
ņé┤ĻĘĀņĀ£ ļ░śņØæ.
ĻĄŁļé┤ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀ 27ĻĘĀņŻ╝ņŚÉ ļīĆĒĢ£ ņé┤ĻĘĀņĀ£ ļ░śņØæņØĆ prochloraz, metconazole, pyraclostrobin, fluxa-pyroxad, dimethomorph, thiophanate-methyl, and copper hydroxideņØś 7ņóģņØä ļīĆņāüņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż(Table 2). ProchlorazļŖö DNA lineage 2ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö Fo080701ĻĘĀņŻ╝ļź╝ ņĀ£ņÖĖĒĢśĻ│ż ņŗ£ļōżņØīļ│æĻĘĀņØś EC50Ļ░ÆņØ┤ 0.02-0.1 ╬╝g/mlļĪ£ ļé«ņØĆ ļåŹļÅäņŚÉņä£ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ĻĘĀņé¼ ņāØņןņØä ņ¢ĄņĀ£ĒĢśņśĆļŗż. MetconazoleņØś EC50Ļ░ÆļÅä 0.04-0.22 ╬╝g/mlļĪ£ prochlorazņÖĆ ļ╣äņŖĘĒĢ£ ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. PyraclostrobinņØś EC50Ļ░ÆņØĆ 0.23-168.01 ╬╝g/mlļĪ£ ĻĘĀņŻ╝ņŚÉ ļö░ļØ╝ ņ░©ņØ┤Ļ░Ć Ēü░ Ļ▓ĮĒ¢źņØä ļ│┤ņśĆņ£╝ļ®░ DNA lineage 1ĻĘĖļŻ╣ņØ┤ ļŗżļźĖ ĻĘĖļŻ╣ņŚÉ ņåŹĒĢśļŖö ĻĘĀļ│┤ļŗż EC50Ļ│╝ EC90Ļ░ÆņØ┤ ĻĘĀņŻ╝ Ļ░ä Ēü░ ņ░©ņØ┤ļź╝ ļ│┤ņśĆļŗż. ĻĘĖ ņÖĖ, fluxapyroxad, dimethomorph, thiophanate-methyl, copper hydroxide 4ņóģņØĆ ļåÆņØĆ ļåŹļÅäņŚÉņä£ļÅä ĻĘĀņé¼ņ¢ĄņĀ£ĒÜ©Ļ│╝Ļ░Ć ļé«ņĢä EC50Ļ░ÆņØä Ļ│äņé░ĒĢĀ ņłś ņŚåņŚłļŗż(Supplementary Fig. 1).
ņ£äņØś Ļ▓░Ļ│╝ļĪ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņŚÉ prochloraz, metconazole, pyraclostrobinņØ┤ ļŗżļźĖ ņé┤ĻĘĀņĀ£ļ│┤ļŗż ĻĘĀņé¼ņ¢ĄņĀ£ĒÜ©Ļ│╝Ļ░Ć ļåÆņĢä ĒÜ©Ļ│╝ņĀüņØĖ ļ░®ņĀ£ņĀ£ļĪ£ņØś Ļ░ĆļŖźņä▒ņØä ļ│┤ņśĆļŗż. ProchlorazļŖö ņĢäņŖżĒīīļØ╝Ļ▒░ņŖżņØś ņżäĻĖ░ņŹ®ņØīļ│æņØä ņØ╝ņ£╝ĒéżļŖö F. oxysporumņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ļ®░(Shin ļō▒, 2018), Colletotrichum spp.ņŚÉ ņØśĒĢ£ ĒāäņĀĆļ│æņŚÉļÅä ļé«ņØĆ ļåŹļÅäņŚÉņä£ ĒÜ©Ļ│╝ņĀüņØĖ ņé┤ĻĘĀņĀ£ņØ┤ļŗż(Kim ļō▒, 2006). MetconazoleņØĆ Fusarium spp.ņŚÉ ļīĆĒĢ£ EC50Ļ░ÆņØ┤ 0.02-0.04 ╬╝g/mlļĪ£ ņāüļŗ╣Ē׳ ļé«ņØĆ ļåŹļÅäņŚÉņä£ ĻĘĀņé¼ ņ¢ĄņĀ£ĒÜ©Ļ│╝Ļ░Ć ņ׳ļŗżĻ│Ā ĒĢśņśĆļŗż(Burlakoti ļō▒, 2010). ĻĘĖļ¤¼ļéś metconazoleņØĆ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀ ņ¢ĄņĀ£ņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ņ¦Ćļ¦ī ļöĖĻĖ░ ņŗØļ¼╝ņ▓┤ ņāØņ£ĪņØä ņ¢ĄņĀ£ĒĢśļŖö ĒÜ©Ļ│╝Ļ░Ć ņ╗ż ņé¼ņÜ®ņŚÉ ņŻ╝ņØśļź╝ ĒĢ┤ņĢ╝ ĒĢĀ ņé┤ĻĘĀņĀ£ņØ┤ļŗż. ļśÉĒĢ£ ĒŖĖļ”¼ņĢäņĪĖĻ│ä ņé┤ĻĘĀņĀ£ņØĖ tebuconazoleņØĆ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņŚÉ ļīĆĒĢ┤ ņĀĆĒĢŁņä▒ ļ░£ļŗ¼ņØś ļČäĒżĻ░Ć ļŗżņ¢æĒĢ£ ĒŖ╣ņä▒ņØä ļ│┤ņØĖļŗżĻ│Ā ĒĢśņśĆļŗż(Kwon ļō▒, 2020). ProchlorazņÖĆ metconazoleĻ│╝ Ļ░ÖņØĆ ĒŖĖļ”¼ņĢäņĪĖĻ│ä ņé┤ĻĘĀņĀ£ļŖö ņŗØļ¼╝ņØś apoplastļĪ£ ļōżņ¢┤Ļ░Ć ļ¼╝Ļ┤Ć, cuticle, ņäĖĒżļ▓Į, ņäĖĒżĻ░äĻĘ╣ņ£╝ļĪ£ ņØ┤ļÅÖĒĢśļŖö demethylation inhibitor (Arntzen ļō▒, 1970)ļĪ£ ļöĖĻĖ░ ļ┐īļ”¼ļĪ£ ņ╣©ņ×ģĒĢśņŚ¼ ļ¼╝Ļ┤ĆņŚÉņä£ ņ”ØņŗØĒĢśļŖö ņŗ£ļōżņØīļ│æņŚÉ ļŗżļźĖ ņé┤ĻĘĀņĀ£ļ│┤ļŗż ĒÜ©Ļ│╝ņĀüņØĖ ļ░®ņĀ£ņĀ£ļØ╝Ļ│Ā ĒĢ£ļŗż.
PyraclostrobinņØĆ soybeanņØś seedling diseaseļź╝ ņØ╝ņ£╝ĒéżļŖö F. oxysporumņŚÉ 0.04-0.481 ╬╝g/mlņØś EC50Ļ░ÆņØä ļ│┤ņŚ¼ ĒÜ©Ļ│╝ņĀüņØĖ ļ░®ņĀ£ņĀ£ļĪ£ ņäĀļ░£ļÉśņŚłļŗż(Cruz Jimenez ļō▒, 2018). Kim ļō▒(2019)ņØĆ Ļ│ĀņČö ĒāäņĀĆļ│æĻĘĀ C. acutatumņØś pyraclostrobinņŚÉ ļīĆĒĢ£ Ļ░Éņłśņä▒ EC50Ļ░ÆņØĆ 0.5 ╬╝g/ml ņØ┤ĒĢś, ņĀĆĒĢŁņä▒ĻĘĀņØĆ 10 ╬╝g/ml ņØ┤ņāüņ£╝ļĪ£ ņĀĆĒĢŁņä▒Ļ│╝ Ļ░Éņłśņä▒ĻĘĀņØś ņ░©ņØ┤ļź╝ ļÜ£ļĀĘņØ┤ ĻĄ¼ļČäĒĢśņśĆņ£╝ļ®░ cyt bņ£ĀņĀäņ×ÉņØś ņĀÉļÅīņŚ░ļ│ĆņØ┤ ļČĆņ£äļź╝ ņØ┤ņÜ®ĒĢ£ ļīĆļ”Įņ£ĀņĀäņ×É ĒŖ╣ņØ┤ņĀü primerļĪ£ļÅä ņĀĆĒĢŁņä▒ĻĘĀņØä ĒīÉļ│äĒĢśņśĆļŗż. PistachioņŚÉ ļ░£ņāØĒĢśļŖö Alternaria alternataņØś pyraclostrobinņŚÉ ļīĆĒĢ£ EC50Ļ░ÆņØĆ Ļ░Éņłśņä▒ <0.01 ╬╝g/ml, ņĢĮņĀĆĒĢŁņä▒ 10 ╬╝g/mlņØ┤ņŚłĻ│Ā(Avenot ļō▒, 2008), ņé¼Ļ│╝ņŚÉ ļ░£ņāØĒĢśļŖö Botrytis cinereaņØś EC50Ļ░ÆņØĆ Ļ░Éņłśņä▒ 0.008-0.132 ╬╝g/ml, ņĀĆĒĢŁņä▒ 0.520-180.3 ╬╝g/mlņØ┤ņŚłļŗż(KimĻ│╝ Xiao, 2010).
ņ£äņÖĆ Ļ░ÖņØĆ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æĻĘĀņŚÉ ļīĆĒĢ£ ņé┤ĻĘĀņĀ£ņØś ĻĘĀņé¼ ņ¢ĄņĀ£ĒÜ©Ļ│╝ ņĪ░ņé¼ļź╝ ĒåĄĒĢ┤ prochlorazņÖĆ Ļ░ÖņØĆ ĒŖĖļ”¼ņĢäņĪĖĻ│╝ pyraclostrobinĻ│╝ Ļ░ÖņØĆ quinol oxidation inhibitorsĻ│äņÖĆ Ēś╝ņÜ®ĒĢśņŚ¼ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņŚÉ ņé¼ņÜ®ĒĢ£ļŗżļ®┤ ļ│┤ļŗż ĒÜ©ņ£©ņĀüņØĖ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢĀ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
ņé┤ĻĘĀņĀ£ņØś ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ĒÜ©Ļ│╝.
ļöĖĻĖ░ ņ£Īļ¼śĒżņןņŚÉņä£ 4ņóģņØś ņé┤ĻĘĀņĀ£ņŚÉ ļīĆĒĢ£ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ņĪ░ņé¼ĒĢ£ Ļ▓░Ļ│╝ boscalid+fludioxonilņØ┤ ļŗżļźĖ ņé┤ĻĘĀņĀ£ļ│┤ļŗż Ļ░Ćņן ĒÜ©Ļ│╝ņĀüņØ┤ņŚłņ£╝ļ®░, fluxapyroxad+pyraclostrobinĻ│╝ prochloraz manganeseļÅä 50% ņØ┤ņāüņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż(Table 3). ļ░śļ®┤ copper hydrox-ideļŖö ņŗ£ļōżņØīļ│æņŚÉ ļé«ņØĆ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż.
ChoņÖĆ Moon (1985)ņØĆ Fof ļ░®ņĀ£ļź╝ ņ£äĒĢ┤ captafol, benomyl, thiophanate-methylņØä ĒåĀņ¢æĻ┤ĆņŻ╝ ņ▓śļ”¼ĒĢśņŚ¼ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. ņĄ£ĻĘ╝ņŚÉļŖö ņ╣£ĒÖśĻ▓Įļ░®ņĀ£ņĀ£ņØĖ sodium dichloroisocyan-urate (NaDCC)ļź╝ ņé¼ņÜ®ĒĢśņŚ¼ 150 ╬╝g/mlņŚÉņä£ FofņØś ĻĘĀņé¼ņāØņןņØ┤ ĒśäņĀĆĒ׳ ņ¢ĄņĀ£ļÉ©ņ£╝ļĪ£ņŹ© ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņĀ£ļĪ£ņØś Ļ░ĆļŖźņä▒ņØ┤ ļ░ØĒśĆņĪīļŗż(Kim ļō▒, 2015). In vitroņāüņŚÉņä£ prochlorazņÖĆ pyraclostrobin
ņØ┤ĻĘĀņé¼ ņāØņן ņ¢ĄņĀ£ĒÜ©Ļ│╝Ļ░Ć ņÜ░ņłśĒ¢łļŖöļŹ░ ņ×¼ļ░░Ēżņן ņŗ£ĒŚśņŚÉņä£ļÅä ņØ┤ļōż ņé┤ĻĘĀņĀ£Ļ░Ć ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ņŚłļŗż. ProchlorazļŖö ĒåĀļ¦łĒåĀņØś ņŗ£ļōżņØīļ│æĻĘĀ F. oxysporum f. sp. lycopersiciņŚÉ ĒÜ©Ļ│╝ņĀüņØĖņĢĮņĀ£ļĪ£ ļ│┤Ļ│ĀļÉśņŚłņ£╝ļ®░(AminiņÖĆ Sidovich, 2010), ĻĄŁļé┤ ļöĖĻĖ░ņ×¼ļ░░ņŚÉ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņĀ£ļĪ£ ļō▒ļĪØļÉśņ¢┤ ņé¼ņÜ®ļÉśĻ│Ā ņ׳ļŗż. ņ£äņØś Ļ▓░Ļ│╝ņŚÉņä£ļŖö boscalid+fludioxonilņØ┤ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ņŚłļŖöļŹ░ĒŖ╣Ē׳ contact ņé┤ĻĘĀņĀ£ņØĖ fludioxonilļ│┤ļŗżļŖö systemic ņé┤ĻĘĀņĀ£ņØĖ boscalidņŚÉ ņØśĒĢ£ ļ░®ņĀ£ĒÜ©Ļ│╝Ļ░Ć Ēü┤ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. FludioxonilņØĆ F. oxysporumņØś ĻĘĀņé¼ ņāØņן ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļŖö ņŚåļŗżĻ│ĀļÅä ĒĢśņśĆļŗż(Cruz Juimenez ļō▒, 2018). Boscalid, fluxapyroxadņÖĆ Ļ░ÖņØĆ SDHIĻ│ä ņé┤ĻĘĀņĀ£ļĪ£ pydiflumetofenņØĆ ņłśļ░ĢņØś ņŗ£ļōżņØīļ│æĻĘĀ(F. oxysporum f. sp. niveum)ņØä ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ļ░®ņĀ£ĒĢ£ļŗżĻ│Ā ĒĢśņśĆļŗż(Miller ļō▒, 2020). ļö░ļØ╝ņä£ ņāłļĪ£ņÜ┤ SDHIĻ│ä ņé┤ĻĘĀņĀ£ņŚÉ ļīĆĒĢ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ Ļ▓ĆĒåĀĒĢ┤ ĒĢäņÜöĻ░Ć ņ׳ņ£╝ļ®░ ļŗżļ¦ī, ņØ┤ļōż ņé┤ĻĘĀņĀ£ļŖö ĻĄÉņ░© ņĀĆĒĢŁņä▒ ļ░£ņāØ Ļ░ĆļŖźņä▒ņØ┤ ļåÆņØĆ ņĢĮņĀ£ļĪ£ ņØ┤ņŚÉ ļīĆĒĢ£ Ļ┤Ćļ”¼ļÅä ĒĢäņÜöĒĢśļŗż.
ņČöĒøä ļöĖĻĖ░ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ļź╝ ņ£äĒĢ£ ņé┤ĻĘĀņĀ£ļŖö FofĻ░Ć ļ┐īļ”¼ ļ░Å Ļ┤ĆļČĆņØś ļÅäĻ┤ĆļČĆņ£äņŚÉņä£ ņ”ØņŗØĒĢśļ»ĆļĪ£ ņé┤ĻĘĀņĀ£Ļ░Ć Ļ┤ĆļČĆņØś ļÅäĻ┤ĆļČĆņ£äĻ╣īņ¦Ć ņ╣©Ēł¼ņØ┤Ē¢ēņØ┤ ļÉśņ¢┤ņĢ╝ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ļ░®ņĀ£ĒĢĀ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ļ│┤ņØ┤ļ®░ ņØ┤ņŚÉ ļīĆĒĢ£ ņé┤ĻĘĀņĀ£ ņäĀļ░£ņØ┤ ņŗ£ĻĖēĒĢ┤ ļ│┤ņØĖļŗż.
ņÜö ņĢĮ
Fusarium oxysporum f. sp. fragariae (Fof) ņŚÉ ņØśĒĢ£ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æņØĆ ĻĄŁļé┤ ļöĖĻĖ░ņ×¼ļ░░ņŚÉņä£ Ļ░Ćņן ņżæņÜöĒĢ£ ļ│æĒĢ┤ ņżæ ĒĢśļéśņØ┤ļŗż. ĻĄŁļé┤ ļ░£ņāØĒĢśļŖö FofņØś ĒŖ╣ņä▒ņØä ļČäņäØĒĢśĻ│Āņ×É ņŗ£ļōżņØīļ│æĻĘĀņØś ņ£ĀņĀäņĀü ļŗżņ¢æņä▒, ļ│æņøÉņä▒Ļ│╝ ņé┤ĻĘĀņĀ£ ļ░śņØæņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ļČäļ”¼ĻĘĀņØĆ Fo080701ļź╝ ņĀ£ņÖĖĒĢ£ ļ¬©ļōĀ ĻĘĀņŻ╝ņŚÉņä£ Fof ĒŖ╣ņØ┤ņĀü primerņŚÉ ņ”ØĒÅŁļÉśņŚłļŗż. ļČäļ”¼ĻĘĀņØś nuclear ribosomal intergenic spacer regionĻ│╝ EF-1 ╬▒ sequences ļČäņäØ Ļ▓░Ļ│╝ 3Ļ░£ņØś lineageļź╝ ĒśĢņä▒ĒĢśņśĆļŗż. ļīĆļČĆļČäņØś ļČäļ”¼ĻĘĀņØĆ lineage 1ņŚÉ ņåŹĒĢśņśĆņ£╝ļ®░ lineage 3ņŚÉ 3Ļ░£ ĻĘĀņŻ╝ņÖĆ lineage 2ņŚÉ 1Ļ░£ ĻĘĀņŻ╝Ļ░Ć ĒżĒĢ©ļÉśņŚłļŗż. ļČäļ”¼ļÉ£ ļ¬©ļōĀ ĻĘĀņŻ╝ļŖö ņäżĒ¢ź ĒÆłņóģņŚÉ ļ│æņøÉņä▒ņØä ļ│┤ņśĆļŗż. ProchlorazļŖö DNA lineage 2ņŚÉ ņåŹĒĢśļŖö Fo080701ĻĘĀņŻ╝ļź╝ ņĀ£ņÖĖĒĢśĻ│ż ņŗ£ļōżņØīļ│æĻĘĀņØś EC50Ļ░ÆņØ┤ 0.02-0.1 ╬╝g/mlļĪ£ ļé«ņØĆ ļåŹļÅäņŚÉņä£ ĒÜ©Ļ│╝ņĀüņ£╝ļĪ£ ĻĘĀņé¼ ņāØņןņØä ņ¢ĄņĀ£ĒĢśņśĆļŗż. MetconazoleņØś EC50Ļ░ÆļÅä 0.04-0.22 ╬╝g/mlļĪ£ prochlorazņÖĆ ļ╣äņŖĘĒĢ£ ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. PyraclostrobinņØś EC50Ļ░ÆņØĆ 0.23-168.01 ╬╝g/mlļĪ£ ĻĘĀņŻ╝ņŚÉ ļö░ļØ╝ ņ░©ņØ┤Ļ░Ć ņ╗Ėļŗż. ļöĖĻĖ░ ņ×¼ļ░░ĒżņןņŚÉņä£ boscalid+fludioxonil, fluxapyroxad+pyraclostrobin, prochloraz manganeseņØ┤ ļöĖĻĖ░ ņŗ£ļōżņØīļ│æ ļ░®ņĀ£ņŚÉ ĒÜ©Ļ│╝ņĀüņØ┤ņŚłļŗż.
Electronic Supplementary Material
Supplementary materials are available at Research in Plant Disease website (http://www.online-rpd.org/).
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Supplement
Supplement Print
Print






