


ņä£ ļĪĀ
ņĮ®(Glycine max Merr.)ņØĆ ļÉ£ņן, Ļ░äņן ļō▒ ņĀäĒåĄņØīņŗØņØś ņøÉļŻīļ┐Éļ¦ī ņĢäļŗłļØ╝, ļŗżņ¢æĒĢ£ ņāØļ”¼ņĀü ĒÜ©Ļ│╝ļź╝ Ļ░Ćņ¦ĆĻ│Ā ņ׳ĻĖ░ ļĢīļ¼ĖņŚÉ ņØśņĢĮ, ņé¼ļŻī, Ļ│ĄņŚģņÜ®Ļ│╝ ņä¼ņ£Ā ļō▒ņØś ņøÉļŻīļĪ£ Ļ┤æļ▓öņ£äĒĢśĻ▓ī ņé¼ņÜ®ļÉśĻ│Ā ņ׳ņ¦Ćļ¦ī, ĻĄŁļé┤ ņĮ® ņ×¼ļ░░ļ®┤ņĀüĻ│╝ ņāØņé░ļ¤ēņØĆ Ļ│äņåŹ Ļ░ÉņåīĒĢśĻ│Ā ņ׳ļŗż. 1981ļģäņŚÉ 201,722haņŚÉņä£ 256,851ĒåżņØ┤ ņāØņé░ļÉśļŗżĻ░Ć, 2000ļģä ļōżņ¢┤ņśżļ®┤ņä£ ņ×¼ļ░░ ļ®┤ ņĀüņØĆ 86,176 haļĪ£, ņāØņé░ļ¤ēņØĆ 113,196Ēåżņ£╝ļĪ£ Ļ░ÉņåīĒĢśņśĆļŗż(Korea Statistical Information Service, 2020). Ļ░Éņåī ņČöņäĖļŖö Ļ│äņåŹļÉśņ¢┤ 2018ļģäņØś ņĮ® ņ×¼ļ░░ļ®┤ņĀüĻ│╝ ņāØņé░ļ¤ēņØĆ 50,638 haņÖĆ 89,410ĒåżĻ╣īņ¦Ć ĒĢśļØĮĒĢśņśĆļŗż. ņØ┤ņ▓śļ¤╝ ņĮ®ņØĆ ņ×¼ļ░░ļ®┤ņĀüĻ│╝ ņāØņé░ļ¤ēņØ┤ Ļ░ÉņåīĒĢśĻ│Ā ņ׳ņØä ļ┐Éļ¦ī ņĢäļŗłļØ╝, ņ×¼ļ░░ ĻĖ░Ļ░ä ņżæņŚÉļŖö ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ņŗØļ¼╝ļ│æņøÉņ▓┤Ļ░Ć ņ╣©ņ×ģĒĢśņŚ¼ ļŗżņ¢æĒĢ£ ļ│æņØ┤ ļ░£ņāØĒĢśļ®░ ņāØņé░ļ¤ēņØ┤ ņ£äĒśæļŗ╣ĒĢśĻ│Ā ņ׳ļŗż. ņĀä ņäĖĻ│ä ņĀüņ£╝ļĪ£ ņĮ®ņØś ņāØņé░ļ¤ēņØä ļ¢©ņ¢┤ļ£©ļ”¼ļŖö ņżæņÜöĒĢ£ ļ│æņ£╝ļĪ£ļŖö ņŗ£ņŖżĒŖĖņäĀņČ®ļ│æ, Rhizoctonia, Pythium ļō▒ņŚÉ ņØśĒĢ£ ņ£Āļ¼śĻĖ░ņØś ļ│æ, MacrophominaņŚÉ ņØśĒĢ£ charcoal rot, FusariumņŚÉ ņØśĒĢ£ sudden death syndrome, SeptoriaņŚÉ ņØśĒĢ£ Ļ░łņāēļ¼┤ļŖ¼ļ│æ ļō▒ņØ┤ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ļŗż (Allen ļō▒, 2017). ĻĄŁļé┤ņŚÉļŖö ņ¦äĻĘĀņØ┤ ņØ╝ņ£╝ĒéżļŖö ļ│æņØ┤ ĒāäņĀĆļ│æņØä ļ╣äļĪ» ĒĢśņŚ¼ 24ņóģ, ņäĖĻĘĀņØ┤ ņØ╝ņ£╝ĒéżļŖö ļ│æņØ┤ ļČłļ¦łļ”äļ│æ ļō▒ 4ņóģ, ļ░öņØ┤ļ¤¼ņŖżņŚÉ ņØśĒĢ£ ļ│æĻ│╝ ņäĀņČ®ņŚÉ ņØśĒĢ£ ļ│æņØ┤ Ļ░üĻ░ü 4ņóģĻ│╝ 3ņóģņö®ņØ┤ ļ│┤Ļ│ĀļÉśņ¢┤ ņ׳ ļŗż(Korean Society of Plant Pathology, 2009).
ĻĄŁņĀ£ņĀüņ£╝ļĪ£ļÅä ņĮ®ņØś ņŻ╝ņÜöĒĢ£ ņāØņé░ņ¦ĆņØĖ ļ»ĖĻĄŁĻ│╝ ņ║Éļéśļŗż ņś©ĒāĆļ”¼ņśż ņ¦ĆņŚŁņØś 2010ļģäļČĆĒä░ 2014ļģäĻ╣īņ¦Ć 5ļģäĻ░äņØś ļłäņĀü ņāØņé░ļ¤ēņØĆ 172ņ¢Ą ļČĆņģĖņØĖļŹ░, ļÅÖņØ╝ĒĢ£ ĻĖ░Ļ░ä ļÅÖņĢł ļ│æņ£╝ļĪ£ ņØĖĒĢ£ ņĮ® ņāØņé░ļ¤ē Ļ░ÉņåīļŖö ņĀäņ▓┤ ņāØņé░ļ¤ēņØś 12.5%ņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö 21ņ¢Ą ļČĆņģĖņŚÉ ļŗ¼ĒĢ£ļŗż(Allen ļō▒, 2017). ļ░£ņāØĒĢśļŖö ņŻ╝ņÜö ļ│æņØĆ ņ¦ĆņŚŁņŚÉ ļö░ļØ╝ņä£ ņ░©ņØ┤Ļ░Ć ņ׳ļŖöļŹ░, ļ»ĖĻĄŁ ļČüļČĆ ņ¦ĆņŚŁņØĆ ņĮ® ņŗ£ņŖżĒŖĖņäĀņČ®ņŚÉ ņØśĒĢ£ Ēö╝ĒĢ┤Ļ░Ć Ļ░Ćņן ņ╗ĖļŹś ļ░śļ®┤ņŚÉ, ļé©ļČĆ ņ¦ĆņŚŁņŚÉņä£ļŖö 2010ļģäļČĆĒä░ 2012ļģäĻ╣īņ¦Ć ņŚ░ 3ļģäĻ░ä charcoal rotņŚÉ ņØśĒĢ£ Ēö╝ĒĢ┤Ļ░Ć Ļ░Ćņן ņ╗Ėļŗż. ļ»ĖĻĄŁ ļé©ļČĆ ņ¦ĆņŚŁņŚÉņä£ ņĮ® ņāØņé░ļ¤ē Ļ░ÉņåīņØś ņŻ╝ļÉ£ ņøÉņØĖņØ┤ ļÉśļŖö charcoalrotņØĆ MacrophominaĻ░Ć ņØ╝ņ£╝ĒéżļŖö ļ│æņ£╝ļĪ£, Ļ│Āņś© Ļ▒┤ņĪ░ĒĢ£ ņ¦ĆņŚŁņŚÉņä£ ļ░£ņāØņØ┤ ņŗ¼ĒĢśļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż. ĻĘĖļ¤░ļŹ░ ņĄ£ĻĘ╝ ņĮ® ņ×¼ļ░░ ĻĖ░Ļ░ä ļÅÖņĢłņØś ĻĄŁļé┤ ĻĖ░ņāü ņāüĒÖ®ņØ┤ Ļ│Āņś©Ļ▒┤ ņĪ░ĒĢ┤ņ¦ĆļŖö Ļ▓ĮĒ¢źņØä ļ│┤ņØ┤Ļ│Ā ņ׳ņ¢┤, ĒżņןņŚÉņä£ ņŻ╝ņÜö ļ│æņØś ļ░£ņāØņØ┤ ļ│ĆĒÖöĒĢĀ ņłś ņ׳ļŗżļŖö ņÜ░ļĀżļź╝ ļé│Ļ│Ā ņ׳ļŗż. 1980ļģäņŚÉ 11.2┬░CņØ┤ņŚłļŹś ņŚ░ĒÅē ĻĘĀ ĻĖ░ņś©ņØ┤ 2019ļģäņŚÉļŖö 13.5┬░CļĪ£ ņāüņŖ╣ĒĢśņśĆņ£╝ļ®░, ņĮ® ņ×¼ļ░░ ĻĖ░Ļ░äņØĖ 5ņøöļČĆĒä░ 10ņøöĻ╣īņ¦Ć 6Ļ░£ņøöĻ░äņØś ĒÅēĻĘĀ Ļ░Ģņłśļ¤ēņØä ļ│┤ņĢäļÅä 2013ļģäļČĆĒä░ 2017ļģäĻ╣īņ¦Ć 5ļģäĻ░äņØĆ Ļ│äņåŹ 150 mm ņØ┤ĒĢśļź╝ ĻĖ░ļĪØĒĢśĻ│Ā ņ׳ļŗż (Korea Meteorological Administration, 2020).
ĻĖ░ņāüņØ┤ Ļ│Āņś© Ļ▒┤ņĪ░ĒĢ┤ņ¦Ćļ®┤ņä£ 2016ļģäņŚÉ Ļ▓ĮĻĖ░ļÅä ĒÖöņä▒Ļ│╝ ņŚ░ņ▓£ņ¦ĆņŚŁņØś ĒżņןņŚÉņä£ ņ×¼ļ░░ĒĢśļŹś ņĮ®ņØ┤ ņŗ¼ĒĢśĻ▓ī ņŗ£ļō£ļŖö ņ”ØņāüņØ┤ ļéśĒāĆļé¼ļŗż. ĻĄŁ ļé┤ņŚÉņä£ ņĮ® ļ¦łļ”ä ņ”ØņāüņØä ņØ╝ņ£╝Ēé©ļŗżĻ│Ā ļ│┤Ļ│ĀļÉ£ ļ│æņ£╝ļĪ£ļŖö Diaporthe ņŚÉ ņØśĒĢ£ ļ»ĖņØ┤ļØ╝ļ│æ, FusariumņŚÉ ņØśĒĢ£ ņŗ£ļōżņØīļ│æ, SclerotiumņŚÉ ņØśĒĢ£ ĒØ░ļ╣äļŗ©ļ│æ ļō▒ņØ┤ ņ׳ņ¦Ćļ¦ī(Korean Society of Plant Pathology, 2009), ņĀä ņäĖĻ│ä ņĮ® ņŻ╝ņÜö ņāØņé░ 10Ļ░£ĻĄŁņØś ņāØņé░ļ¤ē Ļ░ÉņåīņØś ņøÉņØĖ ņżæ ļäż ļ▓łņ¦ĖļĪ£ ĒÅēĻ░ĆļÉĀ ļ¦īĒü╝ Ēü░ Ēö╝ĒĢ┤ļź╝ ņŻ╝ļŖö Macrophomina phaseolinaņŚÉ ņØśĒĢ£ charcoal rotņØĆ ņĢīļĀżņĀĖ ņ׳ņ¦Ć ņĢŖņØĆ ņāüĒā£ņØ┤ļŗż (Wrather ļō▒, 1997). ņØ┤ ļ│æņØĆ ļ│æņøÉĻĘĀņØ┤ ņĮ® ļ┐īļ”¼ļź╝ ņ╣©ņ×ģĒĢ£ Ēøä, ņäĖ ĒżĻ░äĻĘ╣ņØä ĒåĄĒĢ┤ņä£ ņ£ĀĻ┤ĆņåŹ ļČĆņ£äņØś ļ¼╝Ļ┤ĆĻ╣īņ¦Ć ņØ┤ļÅÖĒĢśņŚ¼ ņ£ĀĻ┤ĆņåŹ ņĪ░ņ¦üņØä ĻĖ░Ļ│äņĀüņ£╝ļĪ£ ĒÅÉņćäņŗ£ņ╝£ ņŗ£ļōżņØīĻ│╝ ļ¦łļ”ä ņ”ØņāüņØ┤ ļ░£ņāØĒĢśļŖö ļ│æņØ┤ļŗż(SmithņÖĆ Wyllie, 1999). ļ░£ļ│æ ĒøäĻĖ░Ļ░Ć ļÉśļ®┤ ņØ┤ ļ│æņØś ĒŖ╣ņ¦ĢņĀüņØĖ Ēæ£ņ¦ĢņØĖ ņåīļ”ĮĻĘĀĒĢĄ(microsclerotia)ņØ┤ ļ┐īļ”¼ņÖĆ ņżäĻĖ░ņĪ░ņ¦üņŚÉ ļŗżļ¤ē ĒśĢņä▒ļÉ£ļŗż(Gupta ļō▒, 2012). ņØ┤ ļ│æņøÉĻĘĀņŚÉ Ļ░ÉņŚ╝ļÉ£ ĻĖ░ņŻ╝ļŖö ņ┤łĻĖ░ņŚÉļŖö ļÜ£ļĀĘĒĢ£ ņ”ØņāüņØä ļéśĒāĆļé┤ņ¦Ć ņĢŖņ¦Ćļ¦ī, Ļ│Āņś©Ļ▒┤ņĪ░ĒĢ£ ĒÖśĻ▓ĮņĪ░Ļ▒┤ņØ┤ ņ¦ĆņåŹļÉĀ Ļ▓ĮņÜ░ ņŗ£ļōżņØī ņ”ØņāüĻ│╝ Ļ┤┤ņé¼ņ”ØņāüņØ┤ ņ┤ēņ¦äļÉśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Twizeyimana ļō▒, 2012).
ļ│Ė ņŚ░ĻĄ¼ļŖö 2016ļģä Ļ│Āņś©Ļ▒┤ņĪ░ĒĢ£ ĻĖ░ņāü ņĪ░Ļ▒┤ņŚÉņä£ ņŗ£ļōżņØī ņ”ØņāüņØä ļ│┤ņØĖ ņĮ®ņØä Ļ▓ĮĻĖ░ļÅä ĒÖöņä▒Ļ│╝ ņŚ░ņ▓£ņØś ņĮ® ņ×¼ļ░░ ĒżņןņŚÉņä£ ņ▒äņ¦æĒĢśņŚ¼,ņŗ£ļōżņØī ņ”ØņāüņØś ņøÉņØĖņØ┤ ļÉśļŖö ļ│æņøÉĻĘĀņØä ļČäļ”¼ĒĢśņŚ¼ ņ£ĀņĀäņ×É ņłśņżĆņŚÉņä£ ļÅÖņĀĢĒĢśĻ│Ā, ĻĘĀĒĢÖņĀü ĒŖ╣ņ¦ĢĻ│╝ ļ│æņøÉņä▒ ļō▒ņØä ņĪ░ņé¼ĒĢśņŚ¼ ļ│┤Ļ│ĀĒĢśĻ│Āņ×ÉĒĢ£ļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ļ│æņøÉĻĘĀņØś ļČäļ”¼.
2016ļģä 7ņøöļČĆĒä░ 10ņøöĻ╣īņ¦Ć Ļ▓ĮĻĖ░ļÅä ĒÖöņä▒Ļ│╝ ņŚ░ņ▓£ņ¦ĆņŚŁņŚÉņä£ ņŗ£ļōżņØī ņ”ØņāüņØä ļ│┤ņØ┤ļŖö ņĮ®ņØä ņ▒äņ¦æĒĢśņśĆļŗż. ņłśņ¦æĒĢ£ ļ│æļōĀ ņĮ®ņŚÉņä£ ļ│æņøÉĻĘĀņØä ļČäļ”¼ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ Ļ░ÉņŚ╝ļÉ£ ņĪ░ņ¦üņØä ņלļØ╝ 1%ņ░©ņĢäņŚ╝ņåīņé░ļéśĒŖĖļź©(NaOCl) ņÜ®ņĢĪņ£╝ļĪ£ 1ļČäĻ░ä Ēæ£ļ®┤ ņé┤ĻĘĀĒĢśĻ│Ā ņé┤ĻĘĀņłśļĪ£ 3ĒÜī ņäĖņ▓ÖĒĢśņśĆļŗż. ļ®ĖĻĘĀļÉ£ ņŚ¼Ļ│╝ņ¦ĆņŚÉņä£ ļ¼╝ĻĖ░ļź╝ ņĀ£Ļ▒░ĒĢ£ ļŗżņØī, potato dextrose agar (PDA, Difco, Detroit, MI, USA) ļ░░ņ¦Ćņ£äņŚÉ ņś¼ļĀżļåōĻ│Ā 28┬░CņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ļ│æļōĀ ņĪ░ņ¦üņŚÉņä£ ņ×ÉļØ╝ļéśņś© ĻĘĀņé¼ņØś ņäĀļŗ©ļČĆņŚÉņä£ ĻĘĀņé¼ ņĪ░Ļ░üņØä ļ¢╝ņ¢┤ PDA ņé¼ļ®┤ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢ£ Ēøä 4┬░CņŚÉņä£ ļ│┤Ļ┤ĆĒĢśļ®░ ņŗżĒŚśņŚÉ ņé¼ņÜ®ĒĢśņśĆļŗż. ļśÉĒĢ£ ņĮ® ņØ┤ņÖĖņØśņ×æļ¼╝ ņżæņŚÉņä£ ņ░ĖĻ╣©, Ēīź, ļģ╣ļæÉ, ļĢģņĮ® ļō▒ņŚÉņä£ ņŗ£ļōżņØī ņ”ØņāüņØä ļ│┤ņØ┤ļŖöĻ░£ņ▓┤ļź╝ ņ▒äņ¦æĒĢśĻ│Ā ļČäļ”¼ļÉśļŖö ļ│æņøÉĻĘĀņØä ĒśĢĒā£ņĀüņ£╝ļĪ£ ļÅÖņĀĢĒĢśņśĆļŗż.
ļ│æņøÉĻĘĀņØś ĻĘĀņé¼ ņāØņן ņĪ░ņé¼.
ļ│æņøÉĻĘĀ ļ░░ņ¢æņŚÉ ņĀüĒĢ®ĒĢ£ ļ░░ņ¦Ćļź╝ ņĪ░ņé¼ĒĢśĻĖ░ ņ£äĒĢ┤ņä£ ļČäļ”¼ĒĢ£ M. phaseolina HSW16-141 ĻĘĀņŻ╝ļź╝ 28┬░CņØś PDA ļ░░ņ¦ĆņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä, ĻĘĀņé¼ ņäĀļŗ©ļČĆņŚÉņä£ ņ¦üĻ▓Į 7 mmņØś ĻĘĀņé¼ ņĪ░Ļ░üņØä ļ¢╝ņ¢┤ļé┤ņ¢┤ ņāłļĪ£ņÜ┤ PDA ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśĻ│Ā, ņĀĢĒĢ┤ņ¦ä ņś©ļÅä(20┬░C, 25┬░C, 30┬░C, 32┬░C, 35┬░C, 37┬░C, 40┬░C)ņŚÉņä£ ļ░░ņ¢æĒĢśļ®░ ĻĘĀņ┤ØņØś ņ¦üĻ▓ĮņØä ņĪ░ņé¼ĒĢśņśĆļŗż.
ļ│æņøÉĻĘĀņØś ļČäņāØĒżņ×É Ļ┤Ćņ░░.
ChidambaramĻ│╝ Mathur (1975)ņØś ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ļČäņāØĒżņ×Éļź╝ ĒśĢņä▒ņŗ£ņ╝░ļŗż. ļ│┤ļ”¼ ņ×ÄņØä ņĀüļŗ╣ĒĢ£ Ēü¼ĻĖ░ļĪ£ ņ×ÉļźĖ Ēøä ĒØÉļź┤ļŖö ļ¼╝ņŚÉ ņäĖņ▓ÖĒĢśņŚ¼ Ļ▒┤ņĪ░ņŗ£Ēé© Ēøä, 121┬░CņŚÉņä£ 15ļČäĻ░ä ļ®ĖĻĘĀĒĢśņśĆļŗż. ļ®ĖĻĘĀĒĢ£ ļ│┤ļ”¼ ņ×ÄņØĆ ļ»Ėļ”¼ ņżĆļ╣äĒĢ┤ ļæö 1.5% ļ¼╝ĒĢ£ņ▓£ļ░░ņ¦ĆņŚÉ ņś¼ļ”░ Ēøä, PDA ļ░░ņ¦ĆņŚÉņä£ 48ņŗ£Ļ░ä ļ░░ņ¢æĒĢ£ ļ│æņøÉĻĘĀ ĻĘĀņ┤Ø ņäĀļŗ©ņŚÉņä£ ĻĘĀņé¼ ņĪ░Ļ░üņØä ļ¢╝ņ¢┤ ņĀæņóģĒĢśņśĆļŗż. ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢ£ ļ│┤ļ”¼ ņ×ÄņØĆ 30┬░CņŚÉņä£ 12ņŗ£Ļ░äņö® Ļ┤æ/ņĢö ņĪ░Ļ▒┤ņØä ņ£Āņ¦ĆĒĢśļ®░ 7ņØ╝Ļ░ä ļ│┤Ļ┤ĆĒĢśĻ│Ā,ņ×Ä ņ£äņŚÉ ĒśĢņä▒ļÉ£ ļČäņāØĒżņ×ÉĻ░üĻ│╝ ļČäņāØĒżņ×É ĻĘĖļ”¼Ļ│Ā ņåīļ”ĮĻĘĀĒĢĄ ļō▒ņØäĻ┤Ćņ░░ĒĢśņśĆļŗż.
ļ│æņøÉņä▒ Ļ▓ĆņĀĢ.
ļ│æņøÉĻĘĀņØś ļ│æņøÉņä▒ Ļ▓ĆņĀĢņØĆ Gopala ļō▒(2016)ņØśņŗżĒŚś ļ░®ļ▓ĢņØä Ļ░£ņäĀĒĢśņŚ¼ ņŗżņŗ£ĒĢśņśĆļŗż. M. phaseolina HSW16-141 ĻĘĀņŻ╝ļź╝ 32┬░CņØś PDA ļ░░ņ¦Ć(ņĢöņĪ░Ļ▒┤)ņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ņØ┤ņæż ņŗ£Ļ░£ļź╝ 12 mm (ĻĖĖņØ┤)ļĪ£ ņ×ÉļźĖ Ēøä 121┬░CņŚÉņä£ 30ļČä ļÅÖņĢł 1ņØ╝ Ļ░äĻ▓® ņ£╝ļĪ£ ņ┤Ø 2ĒÜī ļ®ĖĻĘĀĒĢśņśĆļŗż. ņāłļĪŁĻ▓ī ņżĆļ╣äĒĢ£ PDA ļ░░ņ¦Ć(300 ┬Ąg/mlņØś streptomycinņØä ņ▓©Ļ░Ć)ņŚÉ ļ®ĖĻĘĀĒĢ£ ņØ┤ņæżņŗ£Ļ░£ 5Ļ░£ļź╝ ņś¼ļĀżļåōņØĆ ļŗżņØī, M. phaseolina HSW16-141ņØś ĻĘĀņ┤Ø ņäĀļŗ©ņŚÉņä£ ņ¦üĻ▓Į 7 mmņØśĻĘĀņé¼ ņĪ░Ļ░üņØä ņלļØ╝ļé┤ņ¢┤ ĒĢśļéśņØś ĒÄśĒŖĖļ”¼ ņĀæņŗ£ļŗ╣ 7Ļ░£ņö® ņĀæņóģĒĢśņśĆļŗż. ļ│æņøÉĻĘĀņØĆ 32┬░C ņĢöņĪ░Ļ▒┤ņŚÉņä£ 7ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼, ņĀæņóģņøÉņ£╝ļĪ£ ņé¼ņÜ®ĒĢśņśĆņ£╝ļ®░, ļīĆņøÉņĮ®ņØä ņś©ņŗżņŚÉņä£ ņ×¼ļ░░ĒĢśņŚ¼ ļ│æņøÉņä▒ ņŗżĒŚśņŚÉ ņé¼ņÜ® ĒĢśņśĆļŗż. ņ×¼ļ░░ņŚÉ ņé¼ņÜ®ĒĢ£ ņøÉņśłņÜ® ņāüĒåĀļŖö 121┬░CņŚÉņä£ 60ļČäĻ░ä 1ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ ņ┤Ø 2ĒÜī ļ®ĖĻĘĀĒĢ£ Ēøä ĒżĒŖĖ(ņ¦üĻ▓Į, 10 cm; ļåÆņØ┤, 9 cm)ņŚÉ ļŗ┤Ļ│Ā, 2% NaOClņŚÉ 2ļČäĻ░ä Ēæ£ļ®┤ņåīļÅģĒĢ£ ļīĆņøÉņĮ® ņóģņ×Éļź╝ 1ļ”Įņö® ĒīīņóģĒĢśņŚ¼ ņ×¼ļ░░ĒĢśņśĆļŗż. ņĮ®ņØś ņ┤łņāØņ×ÄņØ┤ ņÖäņĀäĒ׳ ņĀäĻ░£ļÉśĻ│Ā ņĀ£1ļ│ĖņŚĮņØ┤ ņČ£ĒśäĒĢś ņśĆņØä ļĢī, ņĀæņóģņøÉņ£╝ļĪ£ ņżĆļ╣äĒĢ£ ņØ┤ņæżņŗ£Ļ░£ļĪ£ ņ¦ĆņĀ£ļČĆĻ░Ć Ļ┤ĆĒåĄĒĢśļÅäļĪØ ņ░öļ¤¼ ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢśĻ│Ā ļ®ĖĻĘĀĒĢ£ ņāüĒåĀļź╝ 1 cm Ļ░Ćļ¤ē ļŹ«ņ¢┤ņŻ╝ņŚłļŗż. ļ¼┤ņ▓śļ”¼ĻĄ¼ļŖö ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢśņ¦Ć ņĢŖņØĆ ļ®ĖĻĘĀĒĢ£ ņØ┤ņæżņŗ£Ļ░£ļź╝ ņé¼ņÜ® ĒĢśņŚ¼ ļÅÖņØ╝ĒĢśĻ▓ī ņŗżĒŚśņØä ņłśĒ¢ēĒĢśņśĆļŗż. ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢ£ ņĮ®ņØĆ 32┬░C ņØś ņāØņןņāü(Ļ┤æ/ņĢö, 12 hr/12 hr; ņŖĄļÅä, 50% ņØ┤ņāü)ņ£╝ļĪ£ ņś«Ļ▓© 30ņØ╝Ļ░ä ņ×¼ļ░░ĒĢśļ®░ ļ░£ļ│æņØä ņ£ĀļÅäĒĢśņśĆļŗż.
Internal transcribed spacer (ITS) ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØ.
Ļ▓ĮĻĖ░ļÅä ĒÖöņä▒Ļ│╝ ņŚ░ņ▓£ņØś ļ│æļōĀ ņĮ®ņŚÉņä£ ļČäļ”¼ĒĢ£ HSW16-141, HSB16-159, HSS16-165ņÖĆ YSW16-175, YSB16-178, YSS16-176 ļō▒ ņ┤Ø 6ĻĘĀņŻ╝ļź╝ņäĀļ░£ĒĢśņŚ¼ ITS ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ņŗżņŗ£ĒĢśņśĆļŗż. NucleoSpin PlantŌģĪ (MACHEREY-NAGEL, Duren, Germany) ĒéżĒŖĖļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ļÅÖĻ▓░Ļ▒┤ņĪ░ņŗ£Ēé© ļ│æņøÉĻĘĀņØś ĻĘĀņé¼ļĪ£ļČĆĒä░ genomic DNAļź╝ ņČöņČ£ ĒĢśĻ│Ā, ITS4 (5'-TCC TCC GCT TAT TGA TAT GC-3')Ļ│╝ ITS5 (5'-GGA AGT AAA AGT CGT AAC AAG G-3') ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ITSņśüņŚŁņØä ņ”ØĒÅŁĒĢśņśĆļŗż. ņ£ĀņĀäņ×ÉņØś ņ”ØĒÅŁņØĆ 95┬░CņŚÉņä£ 4ļČä ņ▓śļ”¼ĒĢśĻ│Ā, 94┬░CņŚÉņä£ ļ│Ćņä▒ Ļ│╝ņĀĢ 30ņ┤ł, 48┬░CņŚÉņä£ Ļ▓░ĒĢ® Ļ│╝ņĀĢ 60ņ┤ł, 72┬░CņŚÉņä£ ņŗĀņן Ļ│╝ņĀĢ 60ņ┤łņØś ņŻ╝ĻĖ░ļź╝ ņ┤Ø 30ĒÜī ņŗżņŗ£ĒĢśņśĆņ£╝ļ®░, ļ¦łņ¦Ćļ¦ē ņŗĀņן Ļ│╝ņĀĢņØĆ 72┬░CņŚÉņä£ 7ļČäĻ░ä ņŗżņŗ£ĒĢśņśĆļŗż. ņ”ØĒÅŁļÉ£ PCR ņé░ļ¼╝ņØĆ 1.5% agarose gelņŚÉņä£ ņĀäĻĖ░ņśüļÅÖĒĢśņŚ¼ ļ░┤ļō£ļź╝ ĒÖĢņØĖĒĢśĻ│Ā, PCR purifica-tion kit (eCube PCR Purification Kit, Phile Korea, Seoul, Korea) ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĀĢņĀ£ĒĢśņśĆļŗż. ņĀĢņĀ£ĒĢ£ DNAļŖö Macrogen (Seoul, Korea)ņŚÉ ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ņØśļó░ĒĢśņśĆņ£╝ļ®░, ļČäņäØļÉ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆnBLAST ĒöäļĪ£ĻĘĖļשņ£╝ļĪ£ GenBankņŚÉ ļō▒ļĪØļÉśņ¢┤ ņ׳ļŖö MacrophominaņåŹ Ļ│░ĒīĪņØ┤ ĻĘĀņŻ╝ņÖĆ ņĮ® ņłśĒÖĢĻĖ░ņŚÉ ļ░£ņāØĒĢśļŖö ņŗØļ¼╝ļ│æņØś ļ│æņøÉĻĘĀ ĻĘĀņŻ╝ņÖĆ ļ╣äĻĄÉĒĢśņśĆļŗż.
ņłśĒÖĢļ¤ēĻ│╝ ĒÆłņ¦ł ņĪ░ņé¼.
ļ░£ļ│æĒżņןņŚÉ ņŗ¼Ļ▓©ņ¦ä ļīĆņøÉņĮ®ņØä ņ£ĪņĢłņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņŚ¼ Ļ▒┤ņĀäĒĢ£ ņĮ®Ļ│╝ ļ│æļōĀ ņĮ®ņ£╝ļĪ£ ĻĄ¼ļ│äĒĢśĻ│Ā, Ļ░üĻ░üņŚÉņä£ ņóģņ×Éļź╝ ņłśĒÖĢĒĢśņśĆļŗż. ņłśĒÖĢĒĢ£ ņóģņ×Éļź╝ ņŗżņś©ņŚÉņä£ 2ņŻ╝ņØ╝Ļ░ä Ļ▒┤ņĪ░ņŗ£Ēé©Ēøä, Ļ▒┤ņĀä ņĮ®Ļ│╝ ļ│æļōĀ ņĮ®ņØś 100ļ”Į ņżæĻ│╝ ņóģņ×ÉņØś ņ¦üĻ▓ĮņØä ņĪ░ņé¼ĒĢśņśĆļŗż.100ļ”Į ņżæņØĆ 10ļ░śļ│Ąņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆĻ│Ā, ņ¦üĻ▓ĮņØĆ 100ļ”ĮņØś ņóģņ×Éļź╝ 3ļ░śļ│Ąņ£╝ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ļ│æņ¦Ģ.
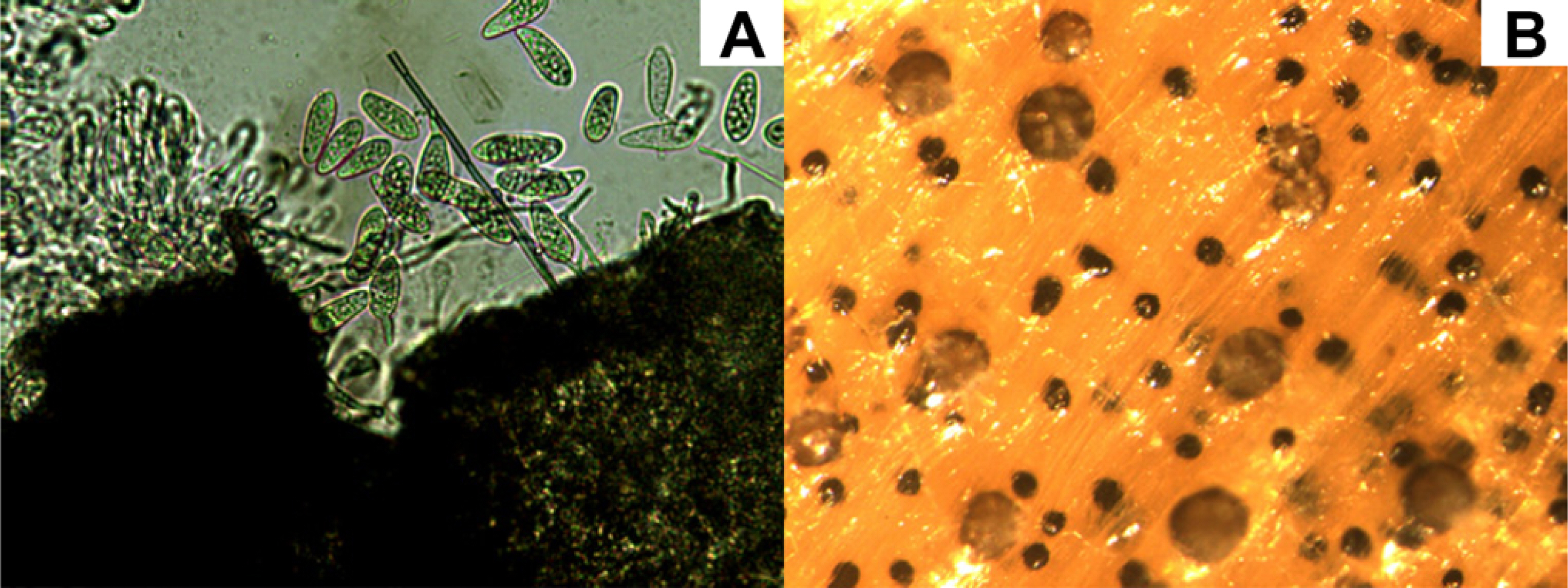
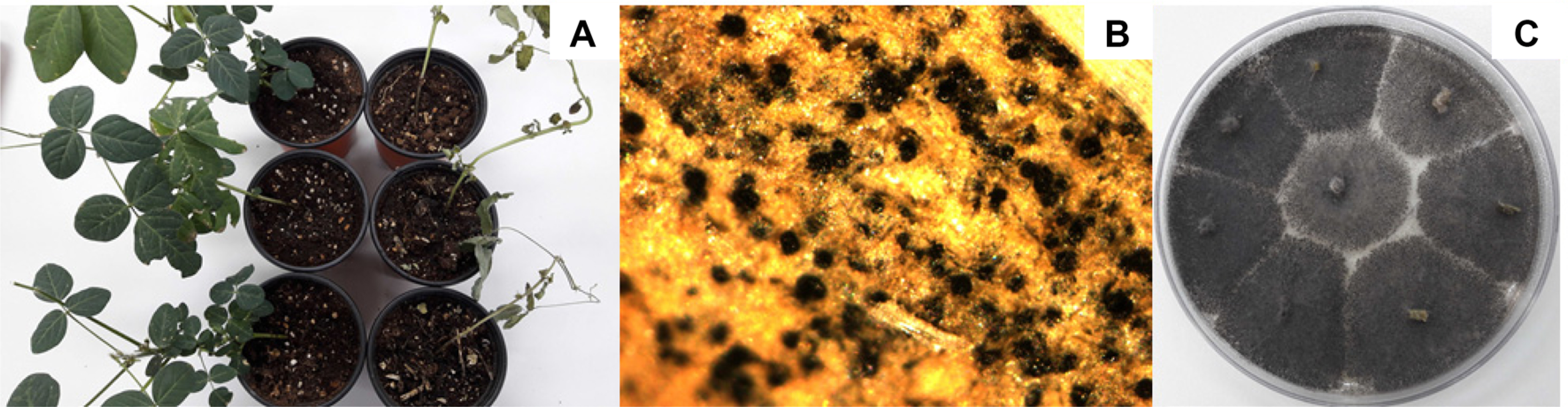
ņĮ® ņ£Āļ¼śĻĖ░ņŚÉ ņ¦ĆņĀ£ļČĆņŚÉņä£ ņłśņ╣©ņāüņØś ļ│æņ¦ĢņØ┤ ĒśĢņä▒ļÉśļŗżĻ░Ć ļ¬©ņלļĪØ ņ”Øņāüņ£╝ļĪ£ ļ░£ņĀäĒĢśņśĆņ£╝ļ®░, ņČ£ņĢäĒĢ£ ļ¢Īņ×ÄņŚÉ Ļ▓ĆņØĆ ļ░śņĀÉņØ┤ļéśĒāĆļéśĻĖ░ļÅä ĒĢśņśĆļŗż(Fig. 1A, B). Ļ░£ĒÖöĻĖ░(2016ļģä 8ņøö 17ņØ╝)ņŚÉ ņ▒äņ¦æĒĢ£ ļ│æļōĀ ņĮ®ņØĆ ņ¦ĆņāüļČĆ ņ×ÄņØ┤ ĒÖ®ĒÖöļÉśĻĖ░ ņŗ£ņ×æĒĢśĻ│Ā ļČĆļČäņĀüņ£╝ļĪ£ Ļ░łļ│ĆĒĢśļŖö ļČĆņ£äļōżņØ┤ ļéśĒāĆļéśĻĖ░ ņŗ£ņ×æĒĢśņśĆļŗż(Fig. 2A). ņ▒äņ¦æĒĢ£ ļ│æļōĀ ņĮ®ņØś ļ┐īļ”¼ ņāØņ£ĪņØĆ ļ¦ÄņØ┤ ņ¢ĄņĀ£ļÉśņ¢┤ ņ׳ņŚłļŖöļŹ░, ņ×öļ┐īļ”¼Ļ░Ć ĒāłļØĮļÉśļ®░ļ┐īļ”¼ ļČĆņ£äņØś Ēæ£Ēö╝Ļ░Ć ļ▓ŚĻ▓©ņ¦Ćļ®┤ņä£ ĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśĻĖ░ ņŗ£ņ×æĒĢśņśĆļŗż(Fig. 2B). Ļ░£ĒÖöĻĖ░ ņØ┤Ēøä 9ņøö 12ņØ╝Ļ▓ĮņŚÉļŖö, ņ¦ĆņāüļČĆņŚÉ ļ¦łļ”ä ņ”ØņāüņØ┤ļÜ£ļĀĘĒĢśĻ▓ī ļéśĒāĆļéśĻĖ░ ņŗ£ņ×æĒĢśņśĆĻ│Ā, ļ┐īļ”¼Ļ│äņØś ņŻ╝ĻĘ╝ņØ┤ ņלļĀżļéśĻ░Ćļ®┤ņä£, Ļ│Āņé¼ĒĢśņśĆļŗż(Fig. 3). ļśÉĒĢ£ ņłśņ╣©ņāüņØś ņ”ØņāüņØä ļ│┤ņØ┤ļŹś ņżäĻĖ░ņŚÉļÅäņåīļ”ĮņØś ĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśņŚłļŗż. ņłśĒÖĢĻĖ░ņØĖ 10ņøö 12ņØ╝ Ļ▓ĮņŚÉ ļ│æņ¦ĢņØä ļ│┤ļ®┤, Fig. 4ņŚÉņä£ ļ│┤ļŖö Ļ▓āĻ│╝ Ļ░ÖņØ┤ ņøÉĻ▒░ļ”¼ņØś ĒżņןņŚÉņä£ļÅä ļ¦łļ”ä ņ”ØņāüņØ┤ ļéśĒāĆļé£ ļČĆņ£äĻ░Ć ļÜ£ļĀĘĒĢśĻ▓ī ĻĄ¼ļ│äļÉśņŚłļŗż. ļśÉĒĢ£ ļ│æļōĀ ņĮ®ņØś ņżäĻĖ░ Ēæ£ļ®┤Ļ│╝ ņ¦ĆņĀ£ļČĆņØś Ēæ£Ēö╝Ļ░Ć ļ▓ŚĻ▓©ņ¦ä ļé┤ļČĆ Ēö╝ņĖĄ ļČĆņ£äņŚÉ ĒśĢņä▒ļÉ£ ĻĘĀĒĢĄņØäļÜ£ļĀĘĒĢśĻ▓ī Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ņŚłļŗż. ļ│æļōĀ Ļ░£ņ▓┤ņØś ņżäĻĖ░ļź╝ ņäĖļĪ£ļĪ£ ņĀłļŗ©ĒĢĀ Ļ▓ĮņÜ░ ņ£ĀĻ┤ĆņåŹ ļČĆņ£äņŚÉ ļŗżļ¤ēņØś ĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉ£ Ļ▓āņØä ļ│╝ ņłś ņ׳ņŚłļŗż (Fig. 4). ņĮ®ņŚÉņä£ņØś ļ│æņ¦ĢņØĆ M. phaseolinaĻ░Ć ļĖöļŻ©ļ▓Āļ”¼ļź╝ ņ╣©ņ×ģĒĢĀ ļĢī ņ¦ĆņāüļČĆņØś ņ×ÄņØ┤ ņŗ£ļōżĻ│Ā, ņżäĻĖ░ņÖĆ ļ┐īļ”¼ļŖö Ļ░łļ│ĆĒĢśļ®┤ņä£ ņŗ£ļō£ļŖö ņ”ØņāüĻ│╝ ņ£Āņé¼ĒĢśņśĆļŗż(de los Santos ļō▒, 2019).Ļ▓░ĻĄŁ, ĒżņןņŚÉņä£ M. phaseolinaĻ░Ć ņĮ®ņØä ņ╣©ņ×ģĒĢśĻ▓ī ļÉśļ®┤ ņĮ®ņ×ÄņØ┤ ņŗ£ļōżļ®┤ņä£ ņĪ░ĻĖ░ņŚÉ ņćĀņĢĮĒĢ┤ņ¦ĆĻ│Ā ņŗØļ¼╝ņ▓┤Ļ░Ć ņØĆĒÜīņāēņ£╝ļĪ£ ļ│ĆņāēļÉśņ¢┤ ņŻĮņŚłļŗż. ņ┤łĻĖ░ņŚÉļŖö ļ┐īļ”¼, ņ¦ĆņĀ£ļČĆ, ņżäĻĖ░ņŚÉ ņłśņ╣©ņāüņØś ņ”ØņāüņØ┤ ļéśĒāĆļéśļ®░, ļ│æņØ┤ ņ¦äņĀäļÉśļ®┤ņä£ ļ┐īļ”¼ņÖĆ ņ¦ĆņĀ£ļČĆņØś Ēæ£Ēö╝ņĪ░ņ¦üņØ┤ ļ▓ŚĻ▓©ņ¦ĆĻ│Ā, ĻĘĖ ņåŹņŚÉ Ļ▓ĆņØĆņāēņØś ņåīļ”ĮĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśĻĖ░ ņŗ£ņ×æĒĢśņśĆļŗż. ņĮ®ņØś ĒÆłņóģņŚÉ ļö░ļØ╝ņä£ Ēæ£Ēö╝Ļ░Ć Ēä░ņ¦Ćļ®┤ņä£ ņä¼ņ£ĀņĪ░ņ¦üņØ┤ Ļ░łļØ╝ņ¦ĆļŖö ņ”ØņāüņØ┤ ļéśĒāĆļéśĻĖ░ļÅä ĒĢśņśĆļŗż. ņØ┤ļ¤¼ĒĢ£ ļ│æņ¦ĢņØä ļ│┤ņØ┤ļŖö ĻĖ░ņŻ╝ļŖö Ļ▒┤ņĀäĒĢ£ ņĮ®Ļ│╝ļŖö ļŗżļź┤Ļ▓ī ļ┐īļ”¼ņÖĆ ņ¦ĆņĀ£ļČĆĻ░Ć ņŹ®Ļ│Ā ņ×öļ┐īļ”¼Ļ░Ć Ļ▒░ņØś ņŚåņ¢┤ ņל ļĮæĒ׳ļ®░, Ļ╝¼Ēł¼ļ”¼Ļ░Ć ņĀ£ļīĆļĪ£ ņŚ¼ļ¼╝ņ¦Ć ņĢŖņĢä ļīĆļČĆļČä Ļ▓░ņŗżņØ┤ ļÉśņ¦Ć ņĢŖņĢśļŗż.
ļ│æņøÉĻĘĀņØś ĻĘĀĒĢÖņĀü ĒŖ╣ņ¦Ģ.
ļ│æļōĀ ņĮ® ņĪ░ņ¦üņŚÉņä£ ļ│æņøÉĻĘĀņØä ļČäļ”¼ĒĢ£ Ļ▓░Ļ│╝, Macrophomina, Phomopsis, Colletotrichum, Botryospaeria, CercosporaņåŹņØś ņŗØļ¼╝ļ│æņøÉĻ│░ĒīĪņØ┤Ļ░Ć ļČäļ”¼ļÉśņŚłņ£╝ļ®░, MacrophominaņØś ļČäļ”¼ļ╣łļÅäļŖö 15.2%ņØ┤ņŚłļŗż(Table 1). ņĮ® ņØ┤ņÖĖņŚÉļÅä ņ░Ė Ļ╣©ņÖĆ ĒīźņŚÉņä£ MacrophominaļŖö Ļ░üĻ░ü 26.7%ņÖĆ 9.1%ņö® ļČäļ”¼ļÉśņŚłļŗż. ĒĢśņ¦Ćļ¦ī ņĮ® ņØ┤ņÖĖņØś ņ×æļ¼╝ņØĆ ņ▒äņ¦æĒĢ£ ņŗ£ļŻīņØś ņłśĻ░Ć ņĀüņŚłļŹś Ļ┤ĆĻ│äļĪ£ ļŹö ņäĖļ░ĆĒĢ£ ņĪ░ņé¼ļź╝ ņŗżņŗ£ĒĢĀ ĒĢäņÜöĻ░Ć ņ׳ļŗż.
ļČäļ”¼ĒĢ£ ļ│æņøÉĻĘĀ M. phaseolina HSW16-141 ĻĘĀņŻ╝ļź╝ 30┬░CņØś PDA ļ░░ņ¦ĆņŚÉņä£ ļ░░ņ¢æĒĢśļ®┤ ĻĘĀņ┤ØņØś ĻĘĀņé¼ ņāēĻ╣öņØ┤ ņ▓śņØīņŚÉļŖö ĒØ░ņāēņØä ļØäļŗżĻ░Ć ļ░░ņ¢æĻĖ░Ļ░äņØ┤ Ļ▓ĮĻ│╝ĒĢśļ®┤ņä£ ĒÜīņāē ļé┤ņ¦Ć Ļ▓ĆņØĆņāēņ£╝ļĪ£ ļ│ĆĒĢśņśĆļŗż. ļ░░ņ¢æĒĢśĻ│Ā 5ņØ╝ņØ┤ ņ¦Ćļéśļ®┤ņä£ļČĆĒä░ PDA ļ░░ņ¦Ć ņāüņŚÉņä£ Ļ▓ĆņĀĢņāēņØś ĻĄ¼ĒśĢņØĖ ņåīļ”ĮĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśĻĖ░ ņŗ£ņ×æĒĢśņśĆņ£╝ļ®░, ĒÅēĻĘĀ ņ¦üĻ▓ĮņØĆ 91 ┬Ąm (ņ¦üĻ▓Į ļ▓öņ£ä, 61-124 ┬Ąm)ņØ┤ņŚłļŗż. ļĖöļŻ©ļ▓Āļ”¼ņØś ļ│æļōĀ ņĪ░ņ¦üņŚÉņä£ ļČäļ”¼ĒĢ£ ļ│æņøÉĻĘĀļÅä 30┬░CņØś ņĢöņāüĒā£ņŚÉņä£ ļ░░ņ¢æĒĢśļ®┤ Ļ▓ĆĻ│Ā ļæźĻĘ╝ ņåīļ”ĮĻĘĀĒĢĄņØä ĒśĢņä▒ĒĢśļŖöļŹ░, ņåīļ”ĮĻĘĀĒĢĄņØś Ēü¼ĻĖ░ļŖö ĒÅēĻĘĀ 98 ┬Ąm (61-128 ┬Ąm)ļĪ£ ņĮ®ņŚÉņä£ ļČäļ”¼ĒĢ£ ņåīļ”ĮĻĘĀĒĢĄĻ│╝ ĒśĢĒā£ņĀüņ£╝ļĪ£ ļ¦żņÜ░ ņ£Āņé¼ĒĢśņśĆļŗż(de los Santos ļō▒, 2019). ļČäņāØĒżņ×ÉņÖĆ ļČäņāØĒżņ×ÉĻ░üņØĆ PDA ļ░░ņ¦ĆņāüņŚÉņä£ļŖö ĒśĢņä▒ļÉśņ¦Ć ņĢŖņĢśļŗż. ĒĢśņ¦Ćļ¦ī ļ®ĖĻĘĀĒĢ£ ļ│┤ļ”¼ ņ×ÄņØä ļ¼╝ĒĢ£ņ▓£ ļ░░ņ¦Ć ņ£äņŚÉ ņś¼ļĀżļåōĻ│Ā ļ│┤ļ”¼ņ×Ä ņ£äņŚÉ M. phaseolina HSW16-141ņØä ņĀæņóģĒĢśņŚ¼ ļ░░ņ¢æĒĢśņśĆņØä ļĢī, Ļ▓ĆņĀĢņāēņØś ļæźĻĘ╝ ļČäņāØĒżņ×ÉĻ░üņØ┤ ĒśĢņä▒ļÉśņŚłņ£╝ļ®░, ļČäņāØĒżņ×ÉĻ░ü ņĢłņŚÉļŖö ļüØņØ┤ ļæźĻĖĆĻ│Ā ĒāĆņøÉĒśĢņØ┤ļ®░ Ēł¼ļ¬ģĒĢ£ ļŗ©Ēżņ×ÉņØĖ ļČäņāØĒżņ×ÉĻ░Ć ļīĆļ¤ēņ£╝ļĪ£ ĒśĢņä▒ļÉśņŚłļŗż(Fig. 5). ļČäņāØĒżņ×ÉĻ░üņØś ĒÅēĻĘĀ ņ¦üĻ▓ĮņØĆ 221 ┬Ąm (157-301 ┬Ąm)ņØ┤ņŚłņ£╝ļ®░, ļČäņāØĒżņ×ÉņØś ņןĻ▓ĮĻ│╝ ļŗ©Ļ▓Į ņØĆ 24 ┬Ąm (16-33 ┬Ąm)ņÖĆ 11 ┬Ąm (9-13 ┬Ąm)ņØ┤ņŚłļŗż. ļ│┤ļ”¼ņ×ÄņŚÉņä£ļÅä Ļ▓ĆņĀĢņāēņØś ĻĄ¼ĒśĢņØĖ ņåīļ”ĮĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśņŚłļŗż.
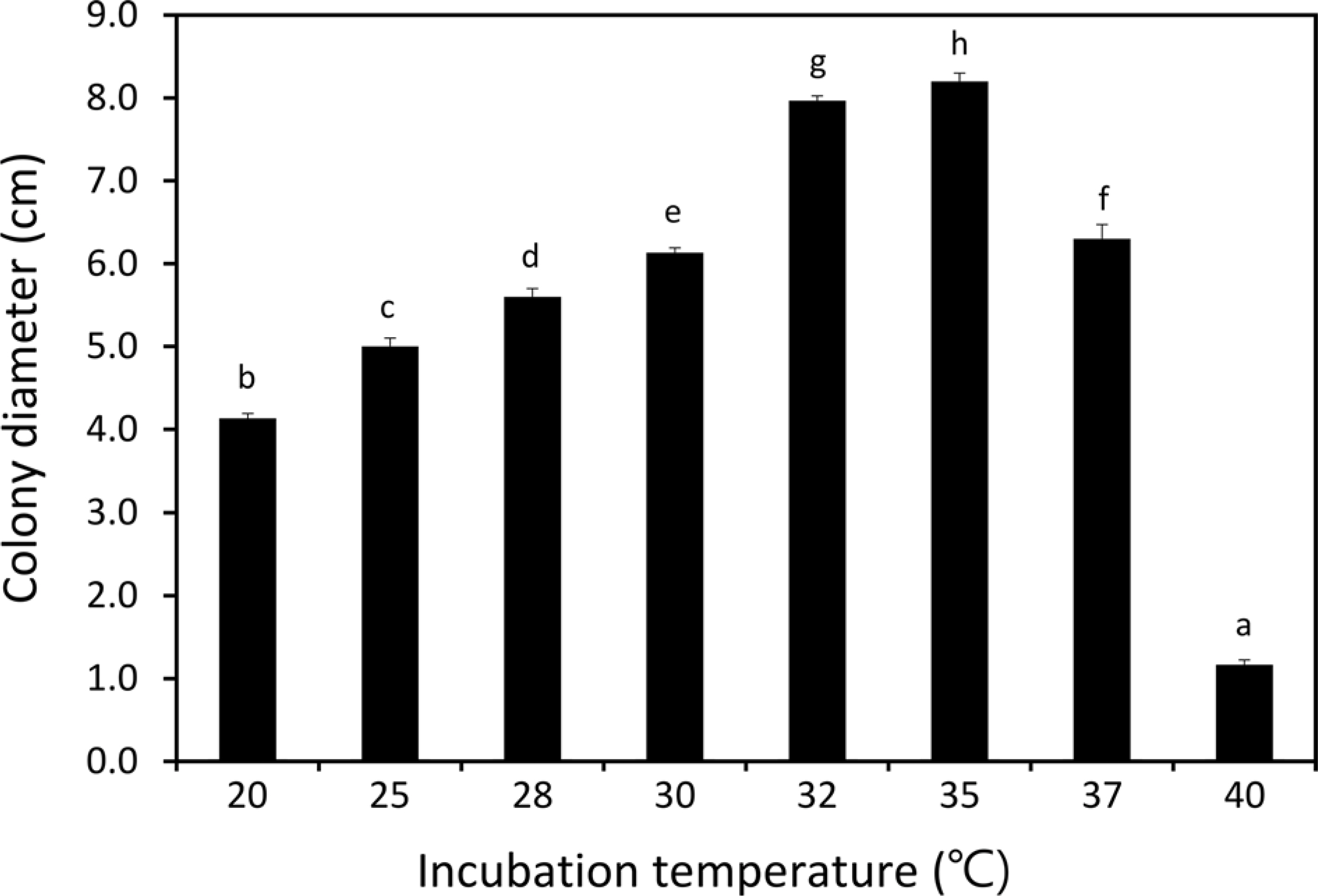
M. phaseolina HSW16-141ņØä PDA ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśĻ│Ā Ļ░üĻ░üņØś ņś©ļÅäņŚÉņä£ 2ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ĻĘĀņ┤ØņØś ņ¦üĻ▓ĮņØä ņĪ░ņé¼ĒĢ£ Ļ▓░Ļ│╝, 35┬░CņŚÉ ņä£ 8.2 cmļĪ£ Ļ░Ćņן ļ╣Āļź┤Ļ▓ī ņāØņןĒĢśņśĆĻ│Ā, 30┬░CņÖĆ 37┬░CņŚÉņä£ļŖö 6.1 Ļ│╝ 6.3 cmļĪ£ ņāØņן ņåŹļÅäĻ░Ć ņĀĆĒĢśļÉśņŚłļŗż. ņØ╝ļ░śņĀüņØĖ ņŗØļ¼╝ļ│æņøÉĻ│░ĒīĪ ņØ┤ņØś ņāØņן ņĀüņś©ņ£╝ļĪ£ ņĢīļĀżņ¦ä 20┬░C, 25┬░C, 28┬░CņŚÉņä£ ĻĘĀņé¼ ņāØņן ņĀĢļÅäļŖö 4.1, 5.0, 5.6 cmļĪ£ ņĀĆņĪ░ĒĢśņśĆļŗż(Fig. 6). ņżæĻĄŁņØś faba beanņŚÉ ņä£ ļČäļ”¼ĒĢ£ M. phaseolina FMP8-1, FMP8-2, FMP8-4ņÖĆ Ļ░ÖņØĆ ĻĘĀņŻ╝ļŖö ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņé¼ņÜ®ĒĢ£ ņĮ®ņŚÉņä£ ļČäļ”¼ĒĢ£ ĻĘĀņŻ╝ņÖĆļŖö ļŗżļź┤Ļ▓ī 28┬░ CņØś PDA ļ░░ņ¦ĆņŚÉņä£ 12ņŗ£Ļ░ä Ļ┤æņØä ņĪ░ņé¼ĒĢśļ®░ ļ░░ņ¢æĒĢśņśĆņØä ļĢī ņāØņן ņØ┤ ļ╣©ļ×Éļŗż(Sun ļō▒, 2019). ņØ┤ļ¤░ Ļ▓░Ļ│╝ļź╝ ļ│┤ļ®┤ ĻĄŁļé┤ņŚÉņä£ ļČäļ”¼ĒĢśļŖö M. phaseolinaņØś ņāØļ”¼ŃåŹņ£ĀņĀäņĀüņØĖ ļŗżņ¢æņä▒ņØä ņĪ░ņé¼ĒĢ┤ņĢ╝ ĒĢĀ ĒĢäņÜöņä▒ņØ┤ ņ׳ļŗżĻ│Ā ņāØĻ░üĒĢ£ļŗż.
ITS ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØ.
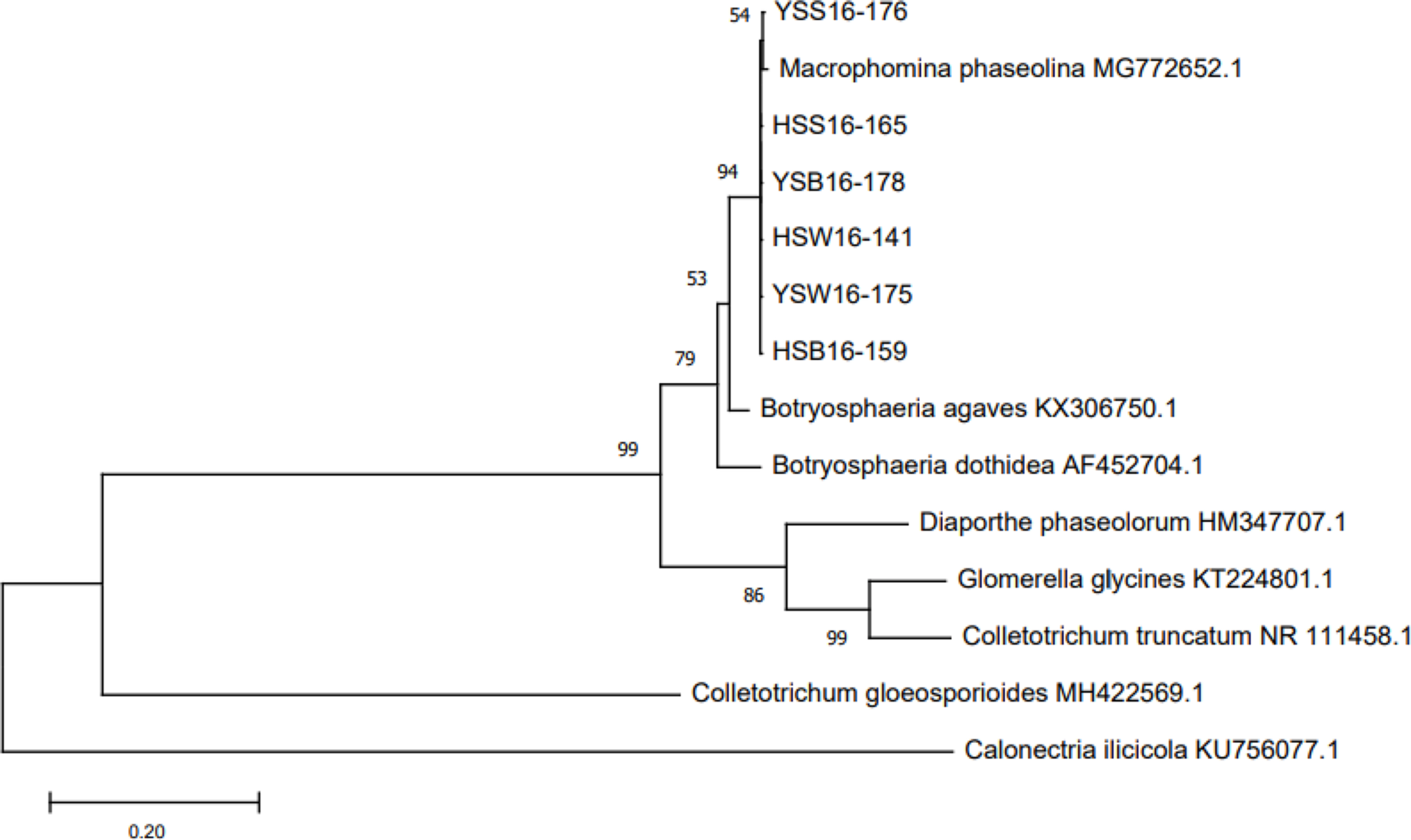
ITS ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ņ£äĒĢ┤ņä£ ITS4ņÖĆ ITS5 ĒöäļØ╝ņØ┤ļ©ĖļĪ£ M. phaseolina ĻĘĀņŻ╝ļōżņØś ņ£ĀņĀäņ×Éļź╝ ņ”ØĒÅŁĒĢśņŚ¼ 580 bpņØś ņ”ØĒÅŁņé░ļ¼╝ņØä ņ¢╗ņŚłļŗż. CummingsņÖĆ Bergstrom (2013) ņØĆ ļ»ĖĻĄŁ ļē┤ņÜĢņŻ╝ņŚÉņä£ ņĮ®ņŚÉ charcoal rot ņ”ØņāüņØä ņØ╝ņ£╝ĒéżļŖö Mp001NY12 ĻĘĀņŻ╝ļĪ£ļČĆĒä░ ITS1Ļ│╝ ITS4 ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ITS ņśüņŚŁņØś ņ£ĀņĀäņ×Éļź╝ ņ”ØĒÅŁĒĢśĻ│Ā ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, M. phaseolinaņÖĆ 100% ņØ╝ņ╣śĒĢśņśĆļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż. Sun ļō▒(2019)ļÅä ņŚŁņŗ£ faba beanņŚÉņä£ ļéśĒāĆļéśļŖö charcoal rotņØś ņøÉņØĖĻĘĀņØä ļČäļ”¼ĒĢśņŚ¼ ņ£ĀņĀäņ×É ņłśņżĆņŚÉņä£ ļÅÖņĀĢņØä ņŗżņŗ£ĒĢśņśĆļŖöļŹ░, ITS4ņÖĆ ITS5 ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņĢĮ 600 bpņØś ITS ņśüņŚŁ ņ£ĀņĀäņ×Éļź╝ ņ”ØĒÅŁĒĢśņśĆņ£╝ļ®░, ņØ┤ ņé░ļ¼╝ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, M. phaseolinaņÖĆ 99-100% ņØ╝ņ╣śĒĢśņśĆļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļÅä ņĮ®ņŚÉņä£ ļČäļ”¼ĒĢ£ 6ĻĘĀņŻ╝ņØś ITS ņśüņŚŁ ņŚ╝ĻĖ░ ņä£ņŚ┤ņØä NCBIņØś GenBankņŚÉ ļō▒ļĪØļÉ£ M. phaseolinaņØś ņŚ╝ĻĖ░ņä£ņŚ┤Ļ│╝ ļ╣äĻĄÉĒĢ£ Ļ▓░Ļ│╝, 100% ņØ╝ņ╣śĒĢśņśĆļŗż. M. phaseolinaņØś ITS ņŚ╝ĻĖ░ņä£ ņŚ┤ņØś Ļ│äĒåĄļČäņäØņØä ĒåĄĒĢ┤ņä£ M. phaseolinaļŖö Botryosphaeriaceae Ļ│╝ņŚÉ ņåŹĒĢśļŖö Botryosphaeria sp.ņÖĆļŖö ĻĘ╝ņŚ░Ļ┤ĆĻ│äņŚÉ ņ׳ņ£╝ļéś, ņĮ® ņłśĒÖĢĻĖ░ņŚÉ ļ░£ņāØĒĢśļŖö Diaporthe, Colletotrichum, Calonectira ļō▒Ļ│╝ ļŖö ļÜ£ļĀĘĒĢ£ ņ░©ņØ┤Ļ░Ć ņ׳ņØīņØä ņĢī ņłś ņ׳ņŚłļŗż(Fig. 7).
ļ│æņøÉņä▒ Ļ▓ĆņĀĢ.
ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢśņŚ¼ ņżĆļ╣äĒĢ£ ņØ┤ņæżņŗ£Ļ░£ļź╝ ņØ┤ņÜ®ĒĢ£ ņāüņ▓ś ņĀæņóģņŚÉņä£ ņĮ®ņØĆ ņĀæņóģĒĢ£ ņ¦Ć 45ņØ╝Ļ▓ĮļČĆĒä░ ļ│æņ¦ĢņØ┤ ļéśĒāĆļé¼ļŗż. ĒżņןņŚÉņä£ Ļ┤Ćņ░░ĒĢ£ ļ░öņÖĆ Ļ░ÖņØ┤ ņĮ®ņØś ņ¦ĆņĀ£ļČĆņŚÉ ņłśņ╣©ņāü ļ│æņ¦ĢņØ┤ ļéśĒāĆļéśļŗżĻ░Ć Ļ░łņāē ļé┤ņ¦Ć Ļ▓ĆņØĆņāēņ£╝ļĪ£ ļ│ĆņāēļÉśĻĖ░ ņŗ£ņ×æĒĢśņśĆņ£╝ļ®░, ļ│æņØ┤ ņ¦äĒ¢ēļÉśļ®┤ņä£ ņĮ®ņØś ņżäĻĖ░ņŚÉņä£ Ļ▓ĆņØĆņāēņØś ņåīļ”ĮĻĘĀĒĢĄņØ┤ ĒśĢņä▒ļÉśņŚłĻ│Ā Ļ▓░ĻĄŁ ņŗ£ļōżņ¢┤ Ļ│Āņé¼ĒĢśņśĆļŗż. ļśÉĒĢ£ ļ│æļōĀ ļČĆņ£äņŚÉņä£ ņĀæņóģĒĢ£ ļ│æņøÉĻĘĀĻ│╝ ļÅÖņØ╝ĒĢ£ M. phaseolinaļź╝ ņ×¼ļČäļ”¼ĒĢĀ ņłś ņ׳ņŚłļŗż(Fig. 8).
ņłśĒÖĢļ¤ēĻ│╝ ĒÆłņ¦ł ņĪ░ņé¼.
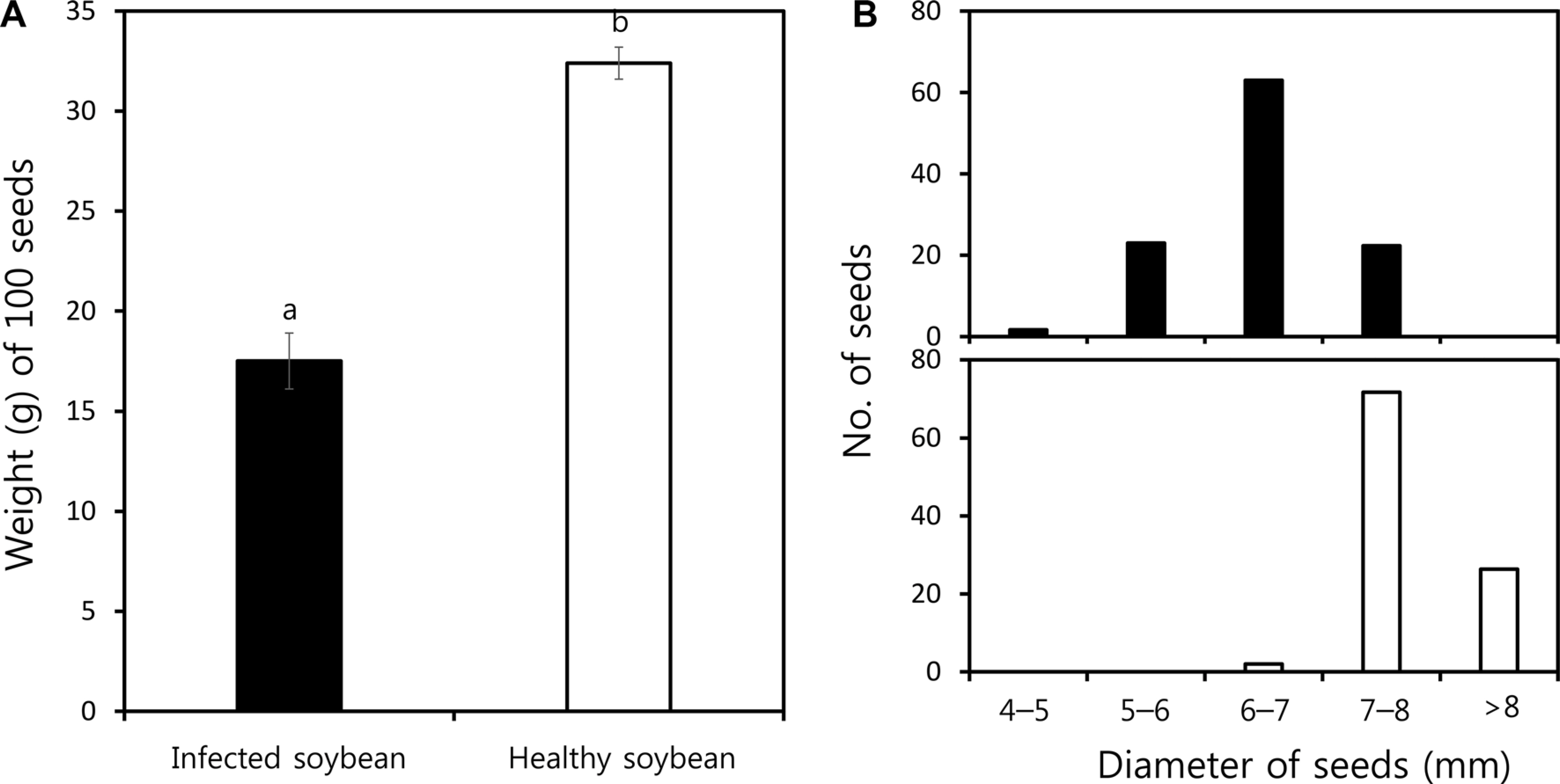
M. phaseolinaĻ░Ć ņ╣©ņ×ģĒĢśņŚ¼ ļ│æņØ┤ ļ░£ņāØĒĢĀ Ļ▓ĮņÜ░ ņĮ®ņØś ņłśĒÖĢļ¤ēĻ│╝ ĒÆłņ¦łņØ┤ ĒśäņĀĆĒĢśĻ▓ī Ļ░ÉņåīĒĢśņśĆļŗż. ļīĆņøÉņĮ® ņØä ņłśĒÖĢĒĢ£ Ēøä, Fig. 9ņŚÉņä£ ļ│┤ļŖö Ļ▓āĻ│╝ Ļ░ÖņØ┤ Ļ▒┤ņĀäĒĢ£ ņĮ® ņóģņ×ÉņÖĆ ļ│æ 10A).ĒĢ┤ņä£ ņĮ® ņóģņ×ÉņØśņ¦üĻ▓ĮņØä ņĪ░ņé¼ĒĢśņśĆļŗż. Ļ▒┤ņĀäĒĢ£ ņĮ® ņóģņ×ÉļŖö ņ¦üĻ▓ĮņØ┤ 8 mm ņØ┤ņāü ņóģņ×ÉĻ░Ć 26%, 7 mm ņØ┤ņāüļČĆĒä░ 8 mm ņØ┤ĒĢśĻ░Ć 72%, 6 mm ņØ┤ņāüņØ┤Ļ│Ā 7mm ņØ┤ĒĢśņØĖ ņóģņ×ÉĻ░Ć 2%ņØĖļŹ░ ļ╣äĒĢśņŚ¼, ļ│æļōĀ ņĮ® ņóģņ×ÉņØś Ļ▓ĮņÜ░ņŚÉļŖö 8mm ņØ┤ņāüņØĖ ņóģņ×ÉļŖö ņŚåĻ│Ā 7 mm ņØ┤ņāüņØ┤ļ®░ 8 mm ņØ┤ĒĢśņØĖ ņóģņ×ÉĻ░Ć12%, 6 mm ņØ┤ņāüņØ┤ļ®░ 7 mm ņØ┤ĒĢśņØĖ Ļ▓āņØ┤ 63%, 5 mm ņØ┤ņāüņØ┤ļ®░6 mm ņØ┤ĒĢśņØĖ Ļ▓āņØ┤ 23%, ĻĘĖļ”¼Ļ│Ā 4 mm ņØ┤ņāüņØ┤ļ®░ 5 mm ņØ┤ĒĢśņØĖ ņóģņ×ÉļÅä 2%ļéś ļÉśņŚłļŗż(Fig. 10B). ņØ┤ņ▓śļ¤╝ M. phaseolinaļŖö ņĮ®ņØś ņāØ ņé░ļ¤ēļ┐Éļ¦ī ņĢäļŗłļØ╝ ņłśĒÖĢĒĢ£ ņĮ®ņØś ĒÆłņ¦łņŚÉļÅä ņśüĒ¢źņØä ļ»Ėņ│żļŗż. ĻĄŁļé┤ņŚÉņä£ļŖö YuņÖĆ Park (1980)ņØ┤ ņ░ĖĻ╣© ņóģņ×ÉņŚÉņä£ ļČäļ”¼ĒĢ£ M. phaseolinaņØś ļ│æņøÉņä▒ņØä ļ│┤Ļ│ĀĒĢśņśĆņ£╝ļ®░, Sung ļō▒(1980)ņØĆ ļ│æļōĀ ņĮ®ņØś ņżäĻĖ░ļéś ļ┐īļ”¼ņŚÉņä£ M. phaseolinaļź╝ ļČäļ”¼ĒĢśĻ│Ā ņĮ®ņŚÉ ļīĆĒĢ£ ļ│æņøÉņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż. YuņÖĆ Park (1982)ņØĆ ĒĢ£ĻĄŁņé░ ņŻ╝ņÜöņ×æļ¼╝ņŚÉņä£ ņóģņ×ÉņĀäņŚ╝ņä▒ ņ¦äĻĘĀņŚÉ ļīĆĒĢ┤ ņĪ░ņé¼ļź╝ ņŗżņŗ£ĒĢśļ®┤ņä£ ņ░ĖĻ╣© ņŗ£ļŻīņŚÉņä£ 0.5-8.5% ļ╣äņ£©ļĪ£ M. phaseolinaļź╝ Ļ▓ĆņČ£ĒĢśņśĆņ£╝ļ®░, YumĻ│╝ Park (1989)ņØĆ Ļ▓ĆņØĆņĮ®ņ£╝ļĪ£ļČĆĒä░ ĒÅēĻĘĀ 0.08%ļĪ£ M. phaseolinaļź╝ Ļ▓ĆņČ£ ĒĢśņśĆļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż. M. phaseolinaļŖö ņóģņ×ÉņĀäņŚ╝Ļ│╝ ĒåĀņ¢æņĀäņŚ╝ņØä ĒĢśĻ│Ā, ĒåĀņ¢æ ņåŹņŚÉņä£ 2-15ļģä ļÅÖņĢł ņāØņĪ┤ņØ┤ Ļ░ĆļŖźĒĢśļ®░(Short ļō▒, 1980), ņåīļ”ĮĻĘĀĒĢĄņØĆ ņØ╝ņ░© ņĀäņŚ╝ņøÉņØś ņŚŁĒĢĀņØä ĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż (Gupta ļō▒, 2012; Kaur ļō▒, 2012). ļö░ļØ╝ņä£ ņĄ£ĻĘ╝ ļōżņ¢┤ ĻĄŁļé┤ ņĮ® ņ×¼ļ░░ ĒżņןņŚÉņä£ ļ░£ņāØĒĢśļŖö M. phaseolinaņŚÉ ņØśĒĢ£ ļ│æņØĆ ļ│æņøÉĻĘĀņØ┤ ņØ┤ļ»Ė ĒåĀņ¢æ ņåŹņŚÉ ņĪ┤ņ×¼ĒĢśĻ│Ā ņ׳ņŚłņ£╝ļ®░, ņĄ£ĻĘ╝ ļ│æņøÉĻĘĀņØś ņāØņןņŚÉ ņĀüĒĢ®ĒĢ£ Ļ│Āņś© Ļ▒┤ņĪ░ĒĢ£ ļéĀņö©Ļ░Ć ņ¦ĆņåŹļÉśļ®┤ņä£ ļ│æņØ┤ ļ░£ņāØĒĢ£ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉśņ¢┤ņ¦äļŗż. ņØ┤ļ»Ė ņśżļל ņĀäņŚÉ YumĻ│╝ Park (1989)ņØ┤ M. phaseolinaņØś ņóģņ×É ņĀäņŚ╝ņØä ļ│┤Ļ│ĀĒĢśņśĆĻ│Ā, ņÜöņ”łņØī ļ│ĆĒÖöĒĢśļŖö ĻĖ░ņāü ņĪ░Ļ▒┤ņ£╝ļĪ£ ņØĖĒĢśņŚ¼ ĒżņןņŚÉņä£ M. phaseolinaņŚÉ ņØśĒĢ£ ļ│æ ļ░£ņāØņØ┤ ņ”ØĻ░ĆĒĢśĻ│Ā ņ׳ļŖö Ļ▓āņØä ļ│┤ļ®┤, ĒżņןņŚÉņä£ ļ░£ņāØ ņ£Āļ¼┤ņÖĆ ļ│æņøÉĻĘĀņØś ļČäĒż, ĻĘĖļ”¼Ļ│Ā ļ│æ ļ░£ņāØ ņāØĒā£ ļō▒ņŚÉ ļīĆĒĢ£ Ē¢źĒøä ņĪ░ņé¼Ļ░Ć ĒĢäņÜöĒĢĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝, ņĮ® ņóģņ×ÉņØś ņāØņé░Ļ│╝ ļ│┤ĻĖē ņĖĪļ®┤ņŚÉņä£ļÅä ļ¦ÄņØĆ ņŻ╝ņØśĻ░Ć ĒĢäņÜöĒĢ£ ņāüĒā£ņØ┤ļŗż.ņØ┤ņāüņØś Ļ▓░Ļ│╝ņŚÉņä£ ļ│┤ļŖö Ļ▓āĻ│╝ Ļ░ÖņØ┤ ņĄ£ĻĘ╝ ĻĖ░Ēøäļ│ĆĒÖöņŚÉ ļö░ļźĖ Ļ│Āņś© Ļ▒┤ņĪ░ĒĢ£ ņĪ░Ļ▒┤ņŚÉņä£ ņĮ®ņŚÉ ļéśĒāĆļéśļŖö ļ¦łļ”ä ņ”ØņāüņØ┤ M. phaseolinaņŚÉ ņØśĒĢ£ Ļ▓āņ×äņØä ļ│┤Ļ│ĀĒĢśļ®┤ņä£, ņĮ®ņŚÉņä£ ļ░£ņāØĒĢ£ M. phaseolinaņŚÉ ņØśĒĢ£ ļ¦łļ”ä ņ”ØņāüņØä ņĮ® ĻĘĀĒĢĄļ¦łļ”äļ│æņ£╝ļĪ£ ļ¬ģļ¬ģĒĢśĻ│Āņ×É ĒĢ£ļŗż.





 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






