

서론
인삼 재배에서 가장 문제가 되는 것은 식물병이다. 그 중에서도 인삼 뿌리에 발생하는 뿌리썩음병으로, Cylindrocarpon destructans와 Fusarium solani에 의한 뿌리썩음병은 인삼 연작장해가 주요 원인으로 여겨지고 있으며, 균핵에 의한 뿌리썩음병, 감자썩이선충에 의한 뿌리썩음병이 발생하고 있다(The Korean Society of Plant Pathology, 2009). 세균에 의해 뿌리에 발생하는 인삼병은 Pectobacterium carotovorum에 의해 발생이 보고되었고(Yu 등, 1991), Paenibacillus polymyxa는 저장 중에 일정한 세균 밀도 이상이 될 때 인삼 뿌리썩음을 유발한다고 하였다(Jeon 등, 2003). Pseudomonas fluorescens와 Serratia liquefaciens는 일정 농도에서 인삼 근부병원균인 Clindrocarpon destructans에 의한 뿌리썩음병 발생정도가 더욱 가속화되도록 하는 역할을 하는 것으로 보고되어 있다(Lee 등, 1975; Kim 등, 2004). 그러나 이들 병원균 단독 감염에 의하여 인삼 뿌리가 부패되는 현상은 많지 않은 것으로 알려져 있다. 2011년 8월 경상북도 봉화군 재산면 농가포장 5년생 인삼포장에서 잎이 초록색을 띈 채 급격하게 시들고 마르는 증상이 발견되었으며 재배하는데 큰 피해를 주고 있었다. 잎이 시든 직후 인삼의 뿌리를 채취하였으나, 뿌리나 줄기에서 특별한 병징은 없었으며, 뿌리의 수분이 약간 빠져나간 상태였다. 이러한 증상이 발생된 지 10일이 경과된 뿌리에서는 뿌리가 물러지면서 썩는 증상이 나타났다. 발병한 인삼뿌리에서 원인균을 찾기 위해 병징 부위에서 병원균을 순수 분리하였다. 그 결과 앞에서 인삼에 보고한 병원세균과 배양 특성이 다르게 나타나 분리 균주의 병원성을 확인하였고 그 병원균을 동정하였다.
병징 및 병원세균의 분리
2011년 8월에 인삼 경작 농업인으로부터 인삼의 잎이 시드는 증상의 원인에 대하여 문의를 받고 경북 봉화의 인삼 재배현장을 방문하였다. 5년생 인삼포장 내 약 10% 정도의 인삼의 잎이 시들었으나, 지상부에 반점이나 물러지는 병징은 발견되지 않았다. 포장에서의 5년생 인삼의 증상은 잎과 줄기가 시들고 잎이 연록색을 띄며 말라 죽는 병징을 나타냈다(Fig. 1). 이러한 증상은 차광망 구조가 잘못되어 빗물이 많이 새는 곳에서 심했다(Fig. 1B). 처음에는 약간 시드는 증상을 나타내고, 인삼을 캐서 보면 잔뿌리 끝 조직이 부패하는 증상을 나타내다가, 후기에는 Fig. 2와 같이 인삼뿌리 끝부분부터 물러지며 썩는 증상을 나타냈다. 잎이 시든 인삼의 뿌리는 급격하게 물러지며 썩어 뿌리의 형태가 사라졌다. 5년생 인삼 포장내 이러한 시들음 증상 발생주율은 10-15%에 달하였으며, 심한 감염주는 뿌리가 모두 부패하여 없어졌다. 잎이 시들고 뿌리가 썩은 인삼을 수집하여 무균상 내에서 인삼의 뿌리 조직을 메스로 잘라내어 차아염소산나트륨 1% 용액과 70% 에탄올(ethyl alcohol)로 표면 살균 후, 멸균수로 세척한 다음 20분 건조하여 Water agar에 올려놓고 배양하였다. 3일 후에 세균 콜로니가 형성되었다. 세균 콜로니를 감자한천배지(PDA)에 streaking하여 단일 콜로니를 분리하였다. 부패한 인삼 뿌리 조직에서 분리된 세균은 PDA 배지에서 흰색의 점질성 콜로니를 형성하며 잘 증식되었다. 분리된 세균은 cryogenic tube에 20% glycerol 용액을 1 ml를 넣어 현탁 후 deep freezer (-70°C)에 보관하며 실험에 사용하였다.


병원성 검정
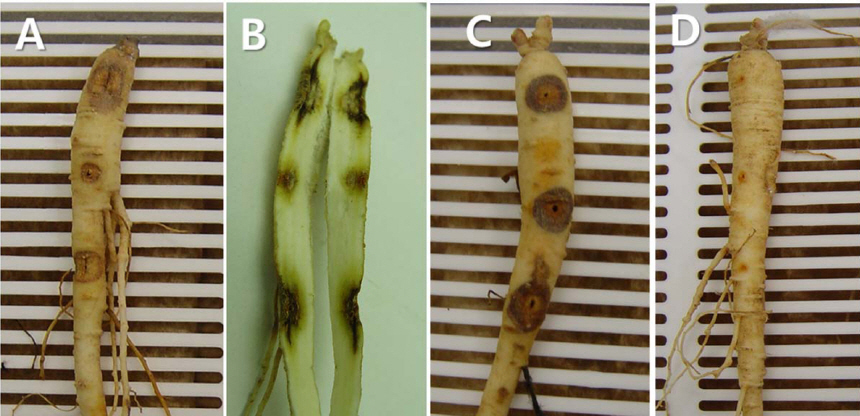
분리된 세균의 인삼에 대한 병원성을 확인하기 위하여 두 가지 방법으로 실험하였다. 첫 번째, 인삼표면에 상처를 내어 세균현탁액을 접종하는 실험을 수행하였다. 인삼은 풍기인삼연구소 포장 내에서 재배되고 있는 2년생 인삼을 이용하였다. 인삼의 표면을 물로 씻고 70% ethyl alcohol로 멸균 후 자연 건조시켰다. PDA에서 2일 동안 배양된 세균을 멸균수에 현탁하여 1×108 CFU/ml 농도로 조정한 다음 5 mm 깊이로 상처 낸 부위에 주사기를 20 μl씩 주입하여 습실 처리한 상자에 넣어 22°C에 보관하며 관찰하였다. 접종 5일 후부터 1×108 CFU/ml 접종 부위가 수침상으로 변하기 시작하였으며, 7일 후부터 접종부위 주변이 갈색으로 변하였고(Fig. 3A, Fig. 3B), 시간이 지남에 따라 암갈색을 띄며 물렁하게 썩기 시작하여 10일 후에는 뿌리가 흑갈색을 띄면서 물러지며 완전히 갈변하였다(Fig. 3C). 병든 부위를 잘라보니 암갈색으로 조직이 변해 있었고, 자른 단면의 암갈색 병반 조직으로부터 처음 접종한 세균과 동일한 세균이 분리되었다. 분리균을 접종하지 않은 음성 대조구에서는 병이 발생하지 않았다(Fig. 3D). 세균 접종 후 2-3주를 두고 관찰하니 세균이 누출되며 인삼뿌리가 완전히 썩었다. 두 번째 실험은 1년생 인삼을 포트에 심고 세균현탁액을 관주하는 방법이다. 묘삼을 3 L 용량의 포트에 5개씩 나누어 심고 1×108 CFU/ml 농도로 조정된 세균현탁액을 500 ml씩 포트에 관주하였다. 처리당 사용된 묘삼은 10개를 사용하였다. 대조구에는 살균수 500 ml을 관주하였다. 1주일에 2회 똑같은 양의 물을 관수하며 실내 15-25°C 온도에서 50일간 병 발생을 조사하였다. 폿트에 묘삼을 심어 세균현탁액을 관주처리한 실험은 10일 후부터 108 CFU/ml 처리구에서 인삼 잎이 시들기 시작하였다. 50일 후 모든 인삼을 뽑아 뿌리의 발병상태를 확인한 결과 세균을 접종한 처리구에서는 병 발생의 차이는 있으나 모두 발병한 것을 확인하였다(Fig. 4A, Fig. 4C). 병원성 검정 후 Koch의 가설을 증명하기 위하여 인삼뿌리에 형성된 병징에서 다시 세균을 분리하였다. 처음 분리된 세균과 다시 분리한 세균의 16S rRNA을 분석하기 위하여 솔젠트 사에 의뢰하여 염기서열 데이터를 받아 NCBI에서 염기서열 유사도를 비교하였고, 동일한 균임을 확인하였다.

병원균의 동정 및 유연관계 분석
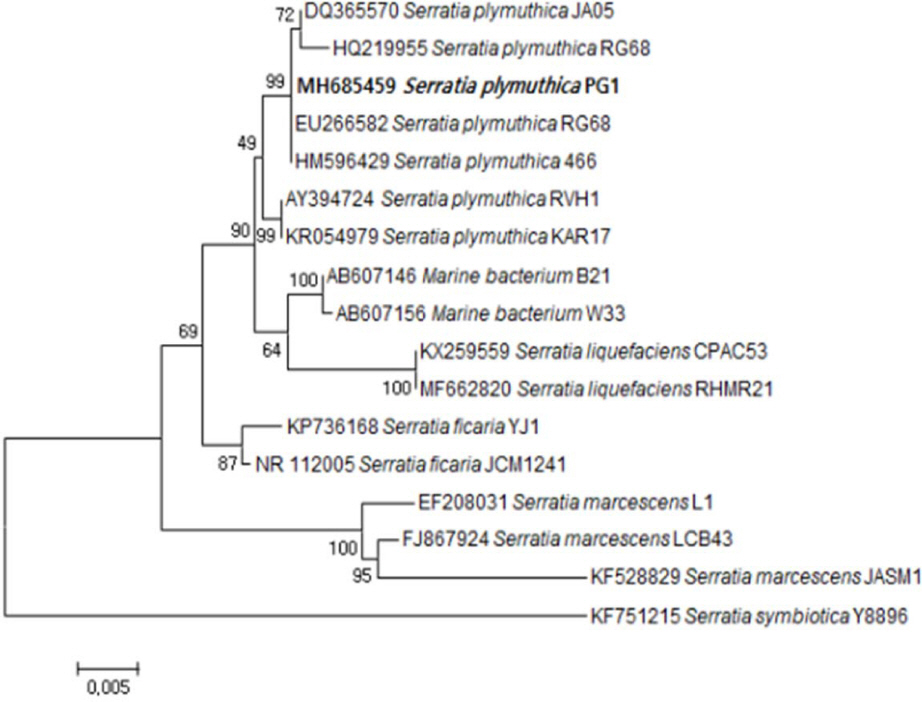
분리균의 탄소원 이용 여부를 조사하기 위하여 TSA (Tryptic Soy Agar)에서 2일 배양한 신선한 콜로니를 면봉으로 채취하여 Inoculating fluid 용액에 현탁하고, Biolog 전용 탁도계를 이용하여 농도를 조절 한 후, GEMIII Microplate (BiologInc., Hayword, Co., USA)에 100 ul씩 분주하여 28°C에서 배양하였다. 24시간 배양 후, MicroLog™3-Automated Microstation System을 이용하여 탄소원 이용 여부를 조사하고, MicroLog Gram-negative database(Version 4.02)와 연결하여 동정하였다. 미생물의 탄소원 및 생화학 특성을 분석하는 Biolog system을 이용하여 동정한 결과 Serratia plymuthica로 85% 유사도, 100%의 가능성으로 동정되었다. 사용하는 탄소원은 Dextrin 등 27개이고 화학물질에 대해서 pH 6 산도에서의 생장, 1% NaCl, 1% Sodium Lactate, Troleandomycin, Rifamycin SV, Lincomycin, Niaproof 4, Vancomycin, Tetrazolium Blue에 양성을 나타내었고, pH 5, 4% NaCl, 8% NaCl, Nalidixic Acid, Fusidic Acid 등에는 음성을 보였다(Table 1). 세균의 생화학적 특성 분석을 위하여 Vitek 2 compact(bioMe’ rieux, France) system을 이용하여 분석하였다. 먼저 TSA에 순수 배양된 세균 콜로니를 5×1 cm 튜브에 0.45% NaCl을 3 ml을 분주한 후 Vitek 전용 탁도계인 Densicheck(bioMe’ rieux, France)를 이용하여 탁도 0.6±0.05 Mcfarland로 현탁하였다. 현탁된 균액은 Vitek 전용 rack에 고정 후 GN card에 현탁액을 충진하였다. 충진부터 이하 일련의 과정은 제조사에서 제공한 실험 방법에 따랐다. Vitek 2 system을 통한 생화학적 반응 결과 S. plymuthica로 99% 동정되었으며, L-Pyrrolydonyl-Arylamidase, β-N-Acetylglucosaminidase, Glutamyl-Arylamidase pNA, γ-Glutamyl Transferase, β-Glucosidase, β-Xylosidase, β-Alanine Arylamidase, Lipase, Sucrose, D-Trehalose에 양성을 보였다(Table 1). 세균의 지방산 분석을 통한 세균 동정 장치인 GC-MIDI를 이용하여 세균을 동정하였다. 세균은 TSA 배지에서 2일간 28°C에서 배양하였다. 세균 콜로니를 1 ml의 saponification reagent (NaOH aqueous methanol)에 첨가하고, 100°C 항온수조에서 30분 간 열을 가한 후, Methylation reagent (hydrochloric acid in aqueous methanol)를 첨가하였다. 지방산은 solvent (hexane/MEBE), mild base (10.8 g NaOH in 900 ml), 그리고 saturated NaOH solution에 의해 추출하였다. 지방산 조성 분석은 Sherlock system을 이용하였으며, MIDI database에 따라 유사도를 측정하였다(MIDI Library version, TSBA 4.0, Library Generation system software version 4.0). 동정 결과 S. plymuthica로 89% 동정되었다. 세균의 16S rDNA 염기서열 분석을 위하여 genomic DNA를 Promega DNA extraction Kit을 이용하여 추출하였고 16S rDNA의 PCR 증폭을 위해 27F (5’-AGAGTTTGATCMTGGCTCAG-3’)와 1492R (5’-GGYTACCTTGTTACGACTT-3’)의 primer를 사용하였다. PCR 반응 조건은 predenaturation(60°C, 2분), denaturation (98°C, 1분), annealing (60°C, 1분), extension (72°C, 1분), total cycle (30 cycle), final extension (72°C, 4분)이었으며, 증폭된 PCR 산물은 1% agarose gel, 0.5 X TBE buffer (0.045 M Tris-borate, 0.001 M EDTA)에서 100 V, 25 mA로 30분 전기영동 후 UV 하에서 확인하였다. PCR 산물을 DNA 염기서열 분석 전문 업체(Solgent, Daejeon, Korea)에 의뢰하였으며, Seqman (DNASTAR, USA) 프로그램을 이용하여 분석하였다. DNASTAR program을 이용하여 염기서열을 붙여 완성한 후 Phylogenetic tree를 작성하고 NCBI (National Center for Biotechnology Information)의 BLAST search를 이용하여 Genbank에 등록되어 있는 균주들과 유사도를 조사하였다. 이세균의 16S rRNA의 염기서열을 분석한 결과, Serratia plymuthica strains와 99.0-99.9%의 homology를 나타냈다(Fig. 5).
Table 1
Comparison of the bacteriological characteristics of the isolates PG1 isolated from decayed ginseng
| Characteristics | Present isolate PG1 | S. plymuthica * | Characteristics | Present isolate PG1 | S. plymuthica * |
|---|---|---|---|---|---|
| Gram stain | - | - | Acid from: | ||
| Indole production | - | - | Fermentation/Glucose | + | + |
| Urease hydrolysis | - | - | β-Glucosidase | + | + |
| Arginine digydrolase | - | - | D-Maltose | - | - |
| Nitrate reduction | + | + | D-Mannitol | + | + |
| Growth in NaCl 1% | + | + | D-Mannose | + | + |
| 4% | - | - | β-Xylosidase | - | - |
| 8% | - | V | β-Alanine arylamidase pNA | - | + |
| Growth in pH5 | - | - | L-Proline Arylamidase | + | + |
| pH6 | + | + | Lipase | - | - |
| Chemical sensitivity: | Palatinose | + | + | ||
| Minocycline | - | - | Tyrosine Arylamidase | + | + |
| Fusidic Acid | - | - | Urease | - | - |
| Rifamycin SV e | + | + | D-Sorbitol | + | V |
| Troleandomycin | + | + | Saccharose/Sucrose | + | + |
| Lincomycin | + | + | D-Tagatose | - | - |
| Guanidine HCl | + | - | D-Trehalose | + | + |
| Niaproof 4 | + | + | Citrate (Sodium) | + | + |
| Vancomycin | + | + | Malonate | - | - |
| Tetrazolium Violet | + | - | 5-Keto-D-Gluconate | + | V |
| Nalidixic Acid | - | - | L-Lactate alkalinisation | + | + |
| Acid from: | α-Glucosidase | + | + | ||
| L-Arabinose | + | + | Sussinate alkalinisation | + | + |
| Glycerol | - | - | β-N-Acetyl-Galactosaminidase | + | + |
| Adonitol | - | - | α-Galactosidase | + | + |
| L-Pyrrolydonyl-Arylamidase | + | + | Phosphatase | + | + |
| L-Arabitor | - | - | Glycine Arylamidase | - | - |
| D-Cellobiose | + | + | Ornithine Decarboxylase | - | - |
| β-Galactosidase | + | + | Lysine Decarboxylase | - | - |
| H2S Production | - | - | Decarboxylase Base | ||
| β-N-Acetyl-Glucosaminidase | + | + | L-Histidine assimilation | - | - |
| Glutamyl Arylamidase pNA | - | - | Coumarate | + | + |
| D-Glucose | + | + | β-Glucoronidase | - | - |
| γ-Glutamyl-Transsferase | + | - | Glu-Gly-Arg-Arylamidase | + | + |
* Details of the Serratia plymuthica are described by Bergey and Holt (1994).
Fig. 5
Phylogenetic relationship between Serratia plymuthica and other Serratia species, constructed using the neighbor-joining tree analysis, based on the combined ITS region and β-tubulin gene sequences. The numbers above the branches represent the bootstrap values obtained for 5,000 replicates.
Serratia 속 세균이 인삼의 뿌리썩음병을 유발시키는 종으로는 Serratia liquefaciens가 보고되었다. Serratia liquefaciens를 인삼 뿌리에 인공 접종하였을 때, 잎은 황화되거나 검은색으로 변하여 시들고 줄기는 속이 썩어 텅 비게 되며, 뿌리는 접종부위가 썩으면서 주름이 생기고 차츰 검은색으로 변한다(Kim 등, 2004). 2011년 폴란드에서 Kowalska 등이 양파 구근에서 이 병원균에 의해 구근썩음병을 일으킨다고 보고 된 바 있다(Kowalska 등, 2011). 따라서 Serratia 속 세균이 인삼의 뿌리썩음 증상을 유발하는 병원성 세균이 존재하는 것으로 보인다. 그러나 Serratia marcescens는 사과 푸른곰팡이병 방제를 위하여 처리하였을 때, 48%의 방제효과가 있다고 보고되었다(Kim 등, 2003). S. plymuthica를 양파 근권에서 분리하여 배양 후, 하우스 내에서 고추 역병에 방제효과가 있는 것으로 보고하였다(Shen 등, 2002, 2005, 2007). S. plymuthica이 생산하는 역병의 포자발아 억제물질은 macrocyclic lactone A21-4으로 밝혀졌다(Shen 등, 2007). 토양 온도가 20°C, 토양 pH가 중성이거나 약산성토양이 S. plymuthica A21-4의 근권정착에 유리하였으며, 살균한 토양보다 토착미생물이 있을 때 토양에 더 잘 정착한다고 하였다(Pu, 2009). 본 연구에서 S. plymutica는 인삼에서 뿌리썩음병을 유발하는 병원균으로 증명되었으며 인삼 세균뿌리썩음병(Bacterial root rot on ginseng)으로 보고한다. 따라서 Serratia plymuthica strain을 인삼에 생물학적 방제제로 적용하는 경우 사용에 주의를 기울여야 할 것이며, 추후 일부 작물에 길항미생물로 경작지에 사용되고 있는 S. plymuthica strain과의 유전적, 생물학적 특성 비교를 수행하여야 할 것으로 생각된다. 이 병은 일반적인 식물세균병과 마찬가지로 토양수분이 과습한 환경에서 다발생하는 것으로 보인다. 차광망 시설에 구멍이 나거나 문제가 있어 빗물이 다량 유입된 두둑 주변의 인삼이 집중적으로 감염되는 것으로 관찰되었다. 차광망을 정상적으로 보수하여 빗물이 누수되지 않도록 하여야 이 병에 의한 피해를 줄일 수 있다.
요약
2011년 8월 경상북도 봉화군 재산면 농가포장 5년생 인삼포장에서 잎이 푸른색을 띈 채 급격하게 마르는 증상이 발견되었다. 잎이 시든 직후 인삼의 뿌리를 채취하였으나, 뿌리나 줄기에서 특별한 병징은 없었으며, 뿌리의 수분이 약간 빠져나간 상태였다. 이러한 증상이 발생된 지 10일이 경과된 뿌리에서는 뿌리가 물러지면서 썩는 증상이 나타났다. 이러한 병징과 건전부의 경계부위에서 세균이 분리되었으며, 이 세균에 의한 뿌리썩음병이 의심되었다. 세균을 배양한 후 건전 인삼에 접종하자, 동일한 썩음 증상이 발현되었으며, 병반으로부터 동일한 세균이 재분리되었다. 이 세균을 BioLog system에 의한 탄소원 이용여부, Vitek 2 system을 이용한 생화학적 반응, GC-MIDI Sherlock system을 이용한 지방산 조성, 16S rRNA 염기서열을 분석한 결과, 모두 Serratia plymuthica로 동정되었다. 이에 따라 인삼 뿌리에 발생하는 이 병을 Serratia plymuthica에 의한 인삼 세균뿌리썩음병으로 명명하여 보고하고자 한다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






