


ņä£ļĪĀ
ĒåĀļ¦łĒåĀļŖö ņ¢æĒīī, ņłśļ░ĢĻ│╝ ĒĢ©Ļ╗ś ņäĖĻ│ä 3ļīĆ ņ▒äņåī ņżæ ĒĢśļéśņØ┤ļŗż. 2015ļģäļÅä ņÜ░ļ”¼ļéśļØ╝ņØś ņĀäņ▓┤ ĒåĀļ¦łĒåĀ ņ×¼ļ░░ļ®┤ņĀüņØĆ 6,975 haļĪ£ ņŻ╝ļĪ£ ņŗ£ņäżņŚÉņä£ ņ×¼ļ░░ļÉśĻ│Ā ņ׳ņ£╝ļ®░, ņČ®ļé©ņØś ņ×¼ļ░░ļ®┤ņĀüņØ┤ 1,604 haļĪ£ Ļ░Ćņן ļäōĻ│Ā, ĻĘĖ ļŗżņØī Ļ░ĢņøÉ, ņĀäļé©ņØś ņł£ņØ┤ļŗż(Ministry of Agriculture, Food and Rural Affairs, 2016). ĒåĀļ¦łĒåĀ ņ×¼ļ░░ ņŗ£ Ēü░ Ēö╝ĒĢ┤ļź╝ ņŻ╝ļŖö ĒÆŗļ¦łļ”äļ│æņØĆ ņŗ£ņäżņ×¼ļ░░ņ¦ĆņØś ņś©ļÅäņÖĆ ņŖĄļÅä ļō▒ņØś ĒÖśĻ▓Į ņÜöņØĖņŚÉ ļö░ļØ╝ ļŗżņ¢æĒĢ£ ļ│æ ļ░£ņāØ ņ¢æņāüņØä ļ│┤ņØĖļŗż(GalleglyņÖĆ Walker, 1949). ĒåĀļ¦łĒåĀļź╝ ĒżĒĢ©ĒĢ£ Ļ░Ćņ¦ĆĻ│╝ ņ×æļ¼╝ņŚÉ ļäōņØĆ ĻĖ░ņŻ╝ ļ▓öņ£äļź╝ Ļ░Ćņ¦ĆĻ│Ā ņ׳ņ£╝ļ®░, ņśżņŚ╝ļÉ£ ĒåĀņ¢æ, ļåŹĻĖ░ĻĄ¼, ļ¼╝Ļ│╝ ĒĢ┤ņČ®ņØä ĒåĄĒĢśņŚ¼ ņĀäĒīīļÉśļ®░, ņłśļģäĻ░ä ĒåĀņ¢æņŚÉ ņāØņĪ┤ĒĢĀ ņłś ņ׳ņ¢┤ņä£ ĒåĀļ¦łĒåĀ ņāØņé░ņŚÉ ņ¦ĆņåŹņĀüņØĖ ņ£äĒśæņÜöņØĖņØ┤ ļÉśĻ│Ā ņ׳ļŗż(TahatņÖĆ Sijam, 2010).
ņŗØļ¼╝ņØĆ ļ│æņøÉĻĘĀļ┐Éļ¦ī ņĢäļŗłļØ╝ Ļ▒┤ņĪ░, Ļ│Āņś©, ņĀĆņś©, ņŚ╝ ļō▒ ļŗżņ¢æĒĢ£ ļ╣äņāØļ¼╝ņĀü ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżļź╝ ļ░øĻ│Ā ņ׳ļŗż(Bohnert ļō▒, 2006; Boyer, 1982). Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖżļĪ£ ņŗØļ¼╝ņØĆ ĒīĮņĢĢņØ┤ ļé«ņĢäņĀĖ Ļ┤æĒĢ®ņä▒ņØ┤ ņĀ£ĒĢ£ļÉśĻ│Ā ņ¦ĆņåŹņĀüņ£╝ļĪ£ Ļ┤æņŚÉ ļģĖņČ£ļÉ©ņ£╝ļĪ£ņŹ© ļ░śņØæņżæņŗ¼ņØ┤ Ļ│╝ļÅäĒĢśĻ▓ī ĒÖśņøÉļÉśņ¢┤ ņŚĮļĪØņ▓┤ņŚÉ ĒÖ£ņä▒ņé░ņåīņóģņØś ņāØņä▒ņØ┤ ņ”ØĻ░ĆĒĢ£ļŗż(Allen, 1995; Smirnoff, 1993). ņŗØļ¼╝ņØĆ 0-15┬░CņØś ņś©ļÅäņŚÉņä£ ņĀĆņś© ņŖżĒŖĖļĀłņŖżļź╝ ļ░øĻ▓ī ļÉśĻ│Ā(Wang, 1989), ņäĖĒżļ¦ēņØś ĻĄ¼ņĪ░ņÖĆ ņĪ░ņä▒ņØś ļ│ĆĒÖö, ņøÉĒśĢņ¦ł ļČäļ”¼ ļō▒ņØś ņäĖĒż ļ│ĆĒÖöļ┐Éļ¦ī ņĢäļŗłļØ╝ ņØ┤ņé░ĒÖöĒāäņåī, ņŚÉĒŗĖļĀīņØś ļ╣äņĀĢņāüņĀüņØĖ ņāØņä▒ ļō▒ ļīĆņé¼Ļ│╝ņĀĢļÅä ļ│ĆĒśĢļÉ£ļŗż. ņĢäņÜĖļ¤¼ ņŗØļ¼╝ņØś ĒÖ£ļĀź ņāüņŗż, ļģĖĒÖö Ļ░ĆņåŹ, ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢ£ Ļ░Éņłśņä▒ņØ┤ ņ”ØĻ░ĆļÉśņ¢┤ ņāØņ£ĪņØ┤ Ļ░ÉņåīļÉśĻ│Ā Ļ▓░ĻĄŁ ņŻĮĻ▓ī ļÉ£ļŗż(SatakeņÖĆ Koike, 1983). ņŗ£ņäżņ×¼ļ░░ņ¦ĆņŚÉņä£ ņŚ╝ļźśņ¦æ ņĀüņØĆ ĒÖöĒĢÖļ╣äļŻīņÖĆ Ēć┤ļ╣äļź╝ Ļ│╝ļÅäĒĢśĻ▓ī ņŗ£ļ╣äĒĢśņŚ¼ ļ░£ņāØĒĢ£ļŗż. ņŚ╝ļźśņ¦æ ņĀüņØĆ ņ×æļ¼╝ņØś ņłśļČä ļ░Å ņ¢æļČä ĒØĪņłśņןĒĢ┤, ņśüņ¢æņןĒĢ┤, ĒÆłņ¦łņĀĆĒĢś ļō▒ņØś ņøÉņØĖņØ┤ ļÉĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņ×æļ¼╝ņØś ļ┐īļ”¼ļź╝ ĻĖ░ĒśĢĒÖöĒĢśņŚ¼ ņŗ£ļōżņØīļ│æĻ│╝ ļ┐īļ”¼Ēś╣ņäĀņČ® ļō▒ņØś Ļ░üņóģ ļ│æĒĢ┤ļź╝ ņØ╝ņ£╝Ēé©ļŗż(Kang ļō▒, 2011).
ĒåĀņ¢æļ»ĖņāØļ¼╝ņØĆ ņ¦üĻ░äņĀæņĀüņ£╝ļĪ£ ņŗØļ¼╝ ņāØņ£ĪņŚÉ ņśüĒ¢źņØä ļ»Ėņ╣śĻ│Ā ņ׳ļŗż(GrossņÖĆ Parthier, 1994; SchrothņÖĆ Hancock, 1982). ņØ┤ ņżæ ņŗØļ¼╝ ņāØņ£Ī ņ┤ēņ¦ä ĻĘ╝ĻČī ņäĖĻĘĀ(plant growth promoting rhizobacteria, PGPR)ņØĆ ņŗØļ¼╝ņØś ļ┐īļ”¼ņé╝ņČ£ņĢĪņŚÉ ņØśĒĢ┤ ņ£ĀļÅäļÉśņ¢┤ ļ¼┤ĻĖ░ņøÉņåīņÖĆ ņłśļČäņØś ĒØĪņłś Ļ░£ņäĀ, ņŗØļ¼╝ĒśĖļź┤ļ¬¼ņØś ļČäļ╣ä ļō▒ņ£╝ļĪ£ ņŗØļ¼╝ņØś ņāØņ£ĪņØä ņ┤ēņ¦äĒĢśĻ│Ā ļ│æņĀĆĒĢŁņä▒Ļ│╝ ņŚ╝, Ļ▒┤ņĪ░ ļō▒ņØś ĒÖśĻ▓ĮņĪ░Ļ▒┤ņŚÉņä£ ļé┤ņä▒ņØä ņ”Øņ¦äņŗ£Ēé©ļŗż(Dodd ļō▒, 2010; GlickmannņÖĆ Dessaux, 1995; TahatņÖĆ Sijam, 2010). PGPRņØĆ ĒÖöĒĢÖļåŹņĢĮ ļ░Å ļ╣äļŻīņŚÉ ļ╣äĒĢ┤ ĒÜ©Ļ│╝ ļ░£ĒśäņØ┤ ļŖ”Ļ│Ā ĒÜ©Ļ│╝Ļ░Ć ļé«ņ£╝ļ®░ ņ¦ĆņåŹĻĖ░Ļ░äņØ┤ ņ¦¦ņĢä ņĀüņÜ®ļ▓öņ£äĻ░Ć ņĀ£ĒĢ£ņĀüņØ┤ļØ╝ļŖö ļŗ©ņĀÉņØ┤ ņ׳ļŗż. ĻĘĖļ¤¼ļéś ņØĖņČĢņŚÉ ļīĆĒĢ£ ļÅģņä▒ņØ┤ ļé«Ļ│Ā, ņāØĒā£Ļ│äņŚÉ ļ»Ėņ╣śļŖö ņśüĒ¢źņØ┤ ņĀüņØĆ ņןņĀÉņØ┤ ņ׳ļŗż. ņĄ£ĻĘ╝ ĒåĀņ¢æĻ░£ļ¤ē, ļ│æĒĢ┤ņČ®ļ░®ņĀ£, ņ£ĀĒĢ┤ļ¼╝ņ¦ł ļČäĒĢ┤, ņ¢æļČäĒØĪņłśņ┤ēņ¦ä, ņ×æļ¼╝ņāØņ£Īņ┤ēņ¦ä, ņĀ£ņ┤ł ļō▒ņØś ņ£ĀņÜ®ĒĢ£ ĻĖ░ļŖźņØä Ļ░Ćņ¦ä PGPRņØä ņäĀļ░£ĒĢśņŚ¼ ļåŹņŚģĒÖśĻ▓Į ļ░Å ņāØĒā£Ļ│äļź╝ ļ│┤ņĪ┤ĒĢśļ®┤ņä£ Ļ│ĀĒÆłņ¦łņØś ļåŹņé░ļ¼╝ņØä ņāØņé░ĒĢśĻĖ░ ņ£äĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ĒÖ£ļ░£Ē׳ ņ¦äĒ¢ēļÉśĻ│Ā ņ׳ļŗż(Hassen ļō▒, 2018; Jung ļō▒, 2007; Mahanty ļō▒, 2017).
ļ│Ė ņŚ░ĻĄ¼ļŖö ĒåĀļ¦łĒåĀ ĻĘ╝ĻČīņŚÉņä£ ņäĖĻĘĀņØä ļČäļ”¼ĒĢśņŚ¼ ņ×æļ¼╝ņØś ņāØņ£Īņ┤ēņ¦ä, ĒÖśĻ▓ĮņŖżĒŖĖļĀłņŖż ļé┤ņä▒ ņ”ØĻ░Ģ ļ░Å ļ│æņøÉĻĘĀ ņ¢ĄņĀ£ļĀźņØä ņ¦Ćļŗī ņ£ĀņÜ® ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśĻ│Ā, ņØ┤ļōż ĻĘĀņŻ╝ņØś Ļ│äĒåĄļČäļźśĒĢÖ ļ░Å ņāØļ”¼ ņāØĒÖöĒĢÖņĀü ĒŖ╣ņä▒ņØä ļČäņäØĒĢśĻ│Āņ×É ĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ĻĘĀņŻ╝ņØś ļČäļ”¼ ļ░Å ļČäļźś
ĒåĀļ¦łĒåĀ ņóģņ×ÉņÖĆ ĻĘ╝ĻČīņŚÉņä£ ĻĘĀņŻ╝ļź╝ ļČäļ”¼ĒĢśĻĖ░ ņ£äĒĢ┤ ņāüĒåĀ(ļČĆļåŹ)ļź╝ Ļ│ĀņĢĢņé┤ĻĘĀĻĖ░ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ 121┬░CņŚÉņä£ 90ļČäĻ░ä 2ĒÜī ņé┤ĻĘĀĒĢśĻ│Ā ņāüņś©ņŚÉņä£ ņŗØĒśöļŗż. ĒåĀļ¦łĒåĀ ņóģņ×É(Solanum lycopersicum, ĒÆłņóģ ņŻ╝ņØ┤ņ╝ä)ļź╝ 70% ņŚÉĒāäņś¼ņŚÉ 1ļČä, 3% ņ░©ņĢäņŚ╝ņåīņé░ļéśĒŖĖļź©ņŚÉ 3ļČäĻ░ä ņé┤ĻĘĀĒĢ£ Ēøä ņé┤ĻĘĀņ”ØļźśņłśļĪ£ 5ĒÜī ņäĖņ▓ÖĒĢśņŚ¼ ņāüĒåĀļź╝ ņČ®ņ¦äĒĢ£ ļ¦łņĀĀĒāĆ ļ░ĢņŖżņŚÉ ĒīīņóģĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀļŖö 28┬░C ņāØņ£ĪņāüņŚÉņä£ 1ņŻ╝ņØ╝Ļ░ä ņāØņ£ĪĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀņØś ļ┐īļ”¼ļź╝ ņĘ©ĒĢśĻ│Ā ĒäĖņ¢┤ļé┤ņ¢┤ ļ╣äĻĘ╝ĻČī ĒåĀņ¢æņØä ņĀ£Ļ▒░ĒĢ£ ļÆż 0.85% NaClņŚÉ ļŗ┤ĻĄ¼ņ¢┤ 15ļČäĻ░ä ņ¦äĒāĢĒĢśņśĆļŗż. 8000 rpmņ£╝ļĪ£ ņøÉņŗ¼ļČäļ”¼ĒĢ£ ĻĘ╝ĻČī ņŗ£ļŻī 1 gņØä 0.85% NaClņŚÉ 10-1Ļ│╝ 10-2ļĪ£ ĒؼņäØĒĢśņŚ¼ ReasonerŌĆÖs 2A (R2A) agar ļ░Å tryptic soy agar (TSA)ņŚÉ ļÅäļ¦É Ēøä 28┬░CņŚÉņä£ 7-10ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ ļŗ©ņØ╝ ņĮ£ļĪ£ļŗłļź╝ ņł£ņłśļČäļ”¼ ĒĢśņśĆļŗż. ņł£ņłśļČäļ”¼ĒĢ£ 56ĻĘĀņŻ╝ļŖö ņé┤ĻĘĀļÉ£ 15% glycerolņŚÉ Ēś╝ĒĢ®ĒĢśņŚ¼ -70┬░CņŚÉ ļ│┤ņĪ┤ĒĢśļ®┤ņä£ ņŗżĒŚśņŚÉ ņØ┤ņÜ®ĒĢśņśĆļŗż. ĻĘĀņŻ╝ņØś ļČäņ×ÉĻ│äĒåĄĒĢÖņĀü ļČäļźśļŖö 16S rRNA ņ£ĀņĀäņ×É ņŗ£ĒĆĆņŖżļź╝ ĻĖ░ļ░śņ£╝ļĪ£ EZ-BioCloud https://www.ezbiocloud.net)ņŚÉ ļō▒ļĪØļÉśņ¢┤ņ׳ļŖö Ēæ£ņżĆ ĻĘĀņŻ╝ņØś ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. ņŚ╝ĻĖ░ņä£ņŚ┤ņØä SINA aligner (http://www.arb-silva.de/aligner)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĀĢļĀ¼ĒĢśĻ│Ā, MEGA6 ĒöäļĪ£ĻĘĖļשņØä ņØ┤ņÜ®ĒĢśņŚ¼ neighbor-joiningļ▓Ģņ£╝ļĪ£ bootstrap 1,000ļ▓ł ļ░śļ│Ą ņŗżĒ¢ēĒĢśņśĆĻ│Ā, Jukes-Cantor modelļĪ£ Ļ│äĒåĄĻ░ä Ļ▒░ļ”¼ļź╝ Ļ│äņé░ĒĢśņśĆļŗż.
ĒåĀļ¦łĒåĀ ņāØņ£Ī ņ┤ēņ¦ä ļ░Å ļ╣äņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ņ”ØĻ░Ģ ĒÜ©Ļ│╝ Ļ▓ĆņĀĢ
ņāØņ£Ī ņ┤ēņ¦ä Ļ▓ĆņĀĢņØĆ 9 cm ļ╣äļŗÉĒżĒŖĖņŚÉ ņāüĒåĀļź╝ ņČ®ņ¦äĒĢśĻ│Ā ĒåĀļ¦łĒåĀļź╝ ĒīīņóģĒĢ£ Ēøä 2ņŻ╝ Ļ░ä ņś©ņŗżņŚÉņä£ ņ×¼ļ░░ĒĢśņśĆļŗż. 1ņŚĮņØ┤ ņĀäĻ░£ļÉśĻĖ░ ņĀä R2Aļ░░ņ¦ĆņŚÉ ņĢĪņ▓┤ļ░░ņ¢æĒĢ£ ĻĘĀņŻ╝ļź╝ 8000 rpmņŚÉ 15ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢśĻ│Ā ņāüļō▒ņĢĪņØä ņĀ£Ļ▒░ĒĢśņśĆļŗż. ņé┤ĻĘĀņ”Øļźśņłśļź╝ ņ▓©Ļ░ĆĒĢśĻ│Ā Ēś╝ĒĢ®ĒĢśņŚ¼ 8000 rpmņŚÉņä£ 15ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ļź╝ 2ĒÜī ļ░śļ│ĄĒĢśņŚ¼ ļ░░ņ¦Ćņä▒ļČäņØä ņĀ£Ļ▒░ĒĢ£ Ēøä 10 mM MgSO4ņŚÉ 5├Ś106 cfu/mlļĪ£ ĒؼņäØĒĢ£ ņäĖĻĘĀĒśäĒāüņĢĪņØä 20 mlņö® Ļ┤ĆņŻ╝ĒĢśĻ│Ā ļ¼┤ņ▓śļ”¼ĻĄ¼ļŖö 10 mM MgSO4ļź╝ ņ▓śļ”¼ĒĢśņśĆļŗż. 4ņŚĮĻĖ░Ļ░Ć ļÉĀ ņŗ£ĻĖ░ņØĖ 2-3ņŻ╝ Ēøä ņ¦ĆņāüļČĆņØś ņāØņ▓┤ņżæņØä ņĖĪņĀĢĒĢśņśĆļŗż. ļ¬©ļōĀ ņŗżĒŚśņØĆ ņ▓śļ”¼ĻĄ¼ļŗ╣ 15ņŻ╝ņØś ļ░śļ│ĄņØä ļæÉņŚłĻ│Ā 3ļ▓łņØś ņŗ£ĻĖ░ņŚÉ Ļ▒Ėņ│É ņŗżņŗ£ĒĢśņśĆņ£╝ļ®░ ļ░░ņ╣śļŖö ņ×äņØśļ░░ņ╣śļź╝ ĒĢśņśĆļŗż.
ņŚ╝ ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ ņ”ØĻ░Ģ Ļ▓ĆņĀĢņØĆ ņāØņ£Īņ┤ēņ¦ä ĒÜ©Ļ│╝ Ļ▓ĆņĀĢĻ│╝ ļÅÖņØ╝ĒĢ£ ļ░®ļ▓Ģņ£╝ļĪ£ Ēīīņóģ Ēøä 2ņŻ╝ņ░©ņŚÉ Yoo ļō▒ (2018)ņØś ļ░®ļ▓ĢņŚÉ ņżĆĒĢśņŚ¼ ņŗ£ĒŚśĻĘĀņŻ╝ļź╝ ĒåĀļ¦łĒåĀ ņ£Āļ¼śņŚÉ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśĻ│Ā 1ņŻ╝ņØ╝ Ēøä Polonenko ļō▒ (1986)ņØś ļ░®ļ▓Ģņ£╝ļĪ£ ņĀ£ņĪ░ĒĢ£ -1000 kPaņØś ļ│ĄĒĢ®ņŚ╝ņÜ®ņĢĪņØä 2ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ 3ĒÜīņŚÉ Ļ▒Ėņ│É 20 mlļź╝ ņ▓śļ”¼ĒĢśĻ│Ā 2ņŻ╝ Ēøä ņāØņ£ĪņØä ņĖĪņĀĢĒĢśņśĆļŗż.
ņś©ļÅä ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ ņ”ØĻ░Ģ Ļ▓ĆņĀĢņØĆ ņāØņ£Ī ņ┤ēņ¦ä ĒÜ©Ļ│╝ Ļ▓ĆņĀĢĻ│╝ ļÅÖņØ╝ĒĢ£ ļ░®ļ▓Ģņ£╝ļĪ£ Ēīīņóģ Ēøä 2ņŻ╝ņ░©ņŚÉ ņŗ£ĒŚśĻĘĀņŻ╝ļź╝ ĒåĀļ¦łĒåĀņŚÉ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśņśĆļŗż. Yoo ļō▒ (2018)ņØś ļ░®ļ▓ĢņØä ļ│ĆĒśĢĒĢśņŚ¼ 1ņŻ╝ņØ╝ Ēøä 10, 25, 40┬░CņØś ņāØņ£ĪņāüņŚÉņä£ 4ņØ╝Ļ░ä ņ£Āņ¦ĆĒĢ£ Ēøä 1ņŻ╝ņØ╝Ļ░ä 25┬░CņØś ĒÖśĻ▓ĮņŚÉņä£ ņł£ĒÖöņŗ£ĒéżĻ│Ā ņāØņ£ĪņØä ņĖĪņĀĢĒĢśņśĆļŗż.
Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ ņ”ØĻ░Ģ Ļ▓ĆņĀĢņØĆ ņāØņ£Ī ņ┤ēņ¦ä ĒÜ©Ļ│╝ Ļ▓ĆņĀĢĻ│╝ ļÅÖņØ╝ĒĢ£ ļ░®ļ▓Ģņ£╝ļĪ£ Ēīīņóģ Ēøä 2ņŻ╝ņ░©ņŚÉ ĻĘĀņŻ╝ļź╝ Ļ┤ĆņŻ╝ņ▓śļ”¼ĒĢśĻ│Ā 1ņŻ╝ ĒøäļČĆĒä░ 15ņØ╝Ļ░ä Ļ┤Ćņłśļź╝ ĒĢśņ¦Ć ņĢŖĻ│Ā Ļ▒┤ņĪ░ĒĢ£ ĒÖśĻ▓ĮņØä ņĪ░ņä▒ĒĢśņśĆļŗż. ĻĘĖ Ēøä 4ņØ╝Ļ░ä ĒåĀļ¦łĒåĀĻ░Ć ĒÜīļ│ĄļÉĀ ņłś ņ׳ļÅäļĪØ Ļ┤ĆņłśĒĢśĻ│Ā ĒåĀļ¦łĒåĀ ņāØņĪ┤ņ£©ņØä ņĖĪņĀĢĒĢśņśĆļŗż. ņāØņĪ┤ņ£©ņØĆ(ņŻĮņØĆ Ļ░£ņ▓┤ņłś/ņ▓śļ”¼ Ļ░£ņ▓┤ņłś)├Ś100%ļĪ£ Ļ│äņé░ĒĢśņśĆļŗż.
ĒåĀļ¦łĒåĀ ĒÆŗļ¦łļ”äļ│æ ņ¢ĄņĀ£ļĀź Ļ▓ĆņĀĢ
9 cm ļ╣äļŗÉ ĒżĒŖĖņŚÉ ĒåĀļ¦łĒåĀļź╝ ĒīīņóģĒĢśĻ│Ā 2ņŻ╝ņ░©ņŚÉ ņŗ£ĒŚśĻĘĀņŻ╝, 10 mM MgSO4(ļīĆņĪ░ĻĄ¼), 0.1 mM benzothiadiazole (BTH)ņØä ĒåĀļ¦łĒåĀņŚÉ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśņśĆļŗż. BTHļŖö ņŗØļ¼╝ņØś ļ®┤ņŚŁļĀźņØä ņ£ĀļÅäĒĢśņŚ¼ ļ│æņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ņØä ņ”ØĻ░Ćņŗ£Ēéżļ»ĆļĪ£ ļ│Ė ņŗżĒŚśņØś ņ¢æņä▒ ļīĆņĪ░ĻĄ¼ļĪ£ ņé¼ņÜ®ĒĢśņśĆļŗż. Ļ┤ĆņŻ╝ņ▓śļ”¼ Ēøä 2ņŻ╝ņØ╝ņØ┤ ņ¦Ćļé£ ņāØņ£Ī 4ņŚĮĻĖ░ņØś ĒåĀļ¦łĒåĀņŚÉ 10 mM MgSO4ļĪ£ ĒśäĒāüĒĢśņŚ¼ OD600=0.25ļĪ£ ņĪ░ņĀłĒĢ£ Ralstonia solanacearum ĻĘĀņŻ╝ ĒśäĒāüņĢĪ 10 mlļź╝ ņĀæņóģĒĢśĻ│Ā 15ņØ╝ņØ┤ Ļ▓ĮĻ│╝ĒĢ£ Ēøä ĒåĀļ¦łĒåĀņØś ļ│æ ļ░£ņāØņ£©ņØä ĒÖĢņØĖĒĢśņśĆļŗż. R. solanacearumņØĆ ĻĄŁļ”ĮļåŹņŚģĻ│╝ĒĢÖņøÉ ļ»ĖņāØļ¼╝ņØĆĒ¢ē(KACC)ņØ┤ ļ│┤ņ£ĀĒĢ£ ĻĘĀņŻ╝(KACC 10695)ļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. ļ│æ ļ░£ņāØļźĀņØĆ (ļ░£ļ│æ Ļ░£ņ▓┤ņłś/ņ▓śļ”¼ Ļ░£ņ▓┤ņłś)├Ś100%ļĪ£ Ļ│äņé░ĒĢśņśĆļŗż. ļ¬©ļōĀ ņŗżĒŚśņØĆ ņ▓śļ”¼ĻĄ¼ļŗ╣ 15ņŻ╝ņØś ļ░śļ│ĄņØä ļæÉņŚłĻ│Ā 3ļ▓łņØś ņŗ£ĻĖ░ņŚÉ Ļ▒Ėņ│É ņŗżņŗ£ĒĢśņśĆņ£╝ļ®░ ļ░░ņ╣śļŖö ņ×äņØśļ░░ņ╣śļź╝ ĒĢśņśĆļŗż.
ĻĘĀņŻ╝ņØś ņāØļ”¼ņāØĒÖöĒĢÖ, ĒśĢĒā£ĒĢÖņĀü ļ░Å ĻĖ░ļŖźņĀü ĒŖ╣ņä▒ ļČäņäØ
ņāØļ”¼ņāØĒÖöĒĢÖ ĒŖ╣ņä▒ ņżæ ņāØņ£Ī ļ░░ņ¦ĆņŗżĒŚśņØĆ ļ»ĖņāØļ¼╝ ĻĘĀņŻ╝ļź╝ R2A, TSA, nutrient agar (NA), potato dextrose agar (PDA) ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 28┬░CņŚÉ 1ņŻ╝ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņāØņ£Ī ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. NaCl ņāØņ£Ī ļ▓öņ£äļŖö ĻĘĀņŻ╝ļź╝ 1-10%(1% Ļ░äĻ▓®)ņØś NaClņØä ņ▓©Ļ░ĆĒĢ£ R2A ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 28┬░CņŚÉ 1ņŻ╝ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņāØņ£Ī ņŚ¼ļČĆļź╝ Ļ┤Ćņ░░ĒĢśņśĆļŗż. ņāØņ£Ī ņś©ļÅä ļ▓öņ£äļŖö ĻĘĀņŻ╝ļź╝ R2A ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 5-50┬░C (5┬░C Ļ░äĻ▓®)ņŚÉņä£ 1ņŻ╝ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņāØņ£Ī ņ£Āļ¼┤ļź╝ Ļ┤Ćņ░░ĒĢśņśĆļŗż. ĒśĢĒā£ĒĢÖņĀü ĒŖ╣ņä▒ņØĆ ņŗżņ▓┤Ēśäļ»ĖĻ▓Į(Carl Zeissņé¼, ļ¬©ļŹĖ DE/MRc5)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ Ļ┤Ćņ░░ĒĢśņśĆļŗż.
PGPR ĒŖ╣ņä▒ ļČäņäØ ņżæ 1-aminocyclopropane-1-carboxylic acid (ACC) Ēāł ņĢäļ»╝ĒÜ©ņåī ĒÖ£ņä▒ņØĆ PenroseņÖĆ Glick (2003) ņØś ļ░®ļ▓Ģņ£╝ļĪ£ DF salt minimal ĒĢ£ņ▓£ļ░░ņ¦Ćļź╝ ņĀ£ņĪ░ĒĢśņŚ¼ Ļ▓ĆņĀĢĒĢśņśĆļŗż. 0.5 MņØś ACC solutionņØĆ 0.2 ╬╝m membrane filterļĪ£ ņŚ¼Ļ│╝ĒĢśņŚ¼ DF salt minimal ļ░░ņ¦ĆņŚÉ 150 ╬╝lņö® ļÅäļ¦ÉĒĢśņśĆļŗż. R2A brothņŚÉņä£ ļ░░ņ¢æļÉ£ ņäĖĻĘĀņØĆ 10 mM MgSO4ņŚÉ ĒśäĒāüĒĢśņŚ¼ OD600=0.25ņØś ļåŹļÅäļĪ£ ļ¦×ņČś Ēøä 100 ╬╝lņö® ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśĻ│Ā 28┬░CņŚÉ 24ņŗ£Ļ░ä ļÅÖņĢł ļ░░ņ¢æĒĢ£ Ēøä ņāØņ£ĪĒ¢łņØä Ļ▓ĮņÜ░ ņ¢æņä▒ņ£╝ļĪ£ ĒīÉņĀĢĒĢśņśĆļŗż. indole-3-acetic acid (IAA) ņāØņä▒ļŖźņØĆ L-tryptophan (100 mg/ml)ņØä ņ▓©Ļ░ĆĒĢ£ R2A brothņŚÉ 28┬░CņŚÉņä£ 1ņØ╝ ļÅÖņĢł ļ░░ņ¢æĒĢ£ Ēøä ņäĖĻĘĀ ļ░░ņ¢æņĢĪņØä 3,000 rpmņ£╝ļĪ£ 20ļČä ļÅÖņĢł ņøÉņŗ¼ļČäļ”¼ĒĢ£ Ēøä ņāüļō▒ņĢĪņØä ļČäņäØņŚÉ ņé¼ņÜ®ĒĢśņśĆļŗż. Salkowski reagentļŖö 50 mlņØś 35% perchloric acidņŚÉ 0.5 M FeCl3 1 mlļź╝ Ēś╝ĒĢ®ĒĢśņŚ¼ ņĀ£ņĪ░ĒĢśņśĆļŗż. ņāüļō▒ņĢĪ 1 mlņÖĆ Salkowski reagent 2 mlļź╝ Ēś╝ĒĢ®ĒĢśņŚ¼ 530 nmņŚÉņä£ ĒØĪĻ┤æļÅäļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. Ēæ£ņżĆ ļåŹļÅäĻ│ĪņäĀņØĆ 0, 1, 2, 4, 8, 10 mMņØś indole-3-acetic acidļź╝ ņé¼ņÜ®ĒĢśņŚ¼ ņ×æņä▒ĒĢśņśĆļŗż(GlickmannņÖĆ Dessaux, 1995; GordonņÖĆ Paleg, 1957; So ļō▒, 2009). ņ¦łņåīĻ│ĀņĀĢļŖźņØĆ Kang ļō▒ (2014)ņØś ļ░®ļ▓ĢņŚÉ ņżĆĒĢśņŚ¼ ņŗ£ĒŚśņÜ® ļ░░ņ¦ĆņŚÉ ĻĘĀņŻ╝ļź╝ ņĀæņóģĒĢ£ Ēøä 28┬░CņŚÉņä£ 3ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņāØņ£ĪĒ¢łņØä Ļ▓ĮņÜ░ ņ¢æņä▒ņ£╝ļĪ£ ĒīÉņĀĢĒĢśņśĆļŗż. ņĢöļ¬©ļŗłņĢä ņāØņä▒ļŖźņØä ņĖĪņĀĢĒĢśĻĖ░ ņ£äĒĢ┤ ĻĘĀņŻ╝ļź╝ ĒÄ®ĒåżņøīĒä░ 10 mlņŚÉ ņĀæņóģĒĢ£ Ēøä 28┬░CņŚÉņä£ 4ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ļ░░ņ¢æņĢĪņØä 8,000 gņŚÉņä£ 15ļČäĻ░ä ņøÉņŗ¼ļČäļ”¼ĒĢ£ Ēøä ņāüļō▒ņĢĪņØä 0.2 ╬╝m membrane filterļĪ£ ņŚ¼Ļ│╝ĒĢśņśĆļŗż. ņŚ¼Ļ│╝ņĢĪ 5 mlņŚÉ Nesseler ņŗ£ņĢĮ 1 mlļź╝ ņ▓©Ļ░ĆĒĢśņŚ¼ ĒÖ®ņāēņŚÉņä£ Ļ░łņāēņ£╝ļĪ£ ļ│ĆņāēļÉśņŚłņØä Ļ▓ĮņÜ░ ņĢöļ¬©ļŗłņĢäņä▒ ņ¦łņåīĻ░Ć ņāØņä▒ļÉ£ Ļ▓āņ£╝ļĪ£ ĒīÉņĀĢĒĢśņśĆļŗż(Dey ļō▒, 2004). ņØĖņé░Ļ░ĆņÜ®ĒÖöļŖźņØĆ Pikovskaya ĒĢ£ņ▓£ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 28┬░CņŚÉ 7ņØ╝Ļ░ä ļ░░ņ¢æ Ēøä ņāØņä▒ļÉ£ Ēł¼ļ¬ģĒÖśņØä ĒÖĢņØĖĒĢśņśĆļŗż(OnyiaņÖĆ Anyanwu, 2013). ņŗ£ĒåĀĒéżļŗī ņāØņä▒ļŖź 0.2% casamino acid, 0.01% thiamine, biotin 2 pgņØ┤ ņ▓©Ļ░ĆļÉ£ M9 ļ░░ņ¦Ć 100 mlņŚÉ ĻĘĀņØä ņĀæņóģĒĢśĻ│Ā 28┬░C, 150 rpmņ£╝ļĪ£ ņ¦äĒāĢ ļ░░ņ¢æĒĢśņśĆļŗż. 72, 96 ļ░Å 120ņŗ£Ļ░äņØ┤ ņ¦Ćļé£ Ēøä 665 nmņŚÉņä£ ĒØĪĻ┤æļÅäļź╝ ņĖĪņĀĢĒĢśņśĆļŗż(PatelņÖĆ Saraf, 2017). ņŗ£ļō£ļĪ£ĒżņĢä ņāØņä▒ļŖźņØĆ CAS diffusion assayļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņĖĪņĀĢĒĢśņśĆļŗż(Shin ļō▒, 2001). ņé┤ĻĘĀļÉ£ ļ®öņŖżļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņŗżĒŚśņÜ® ļ░░ņ¦Ćļź╝ 2ļō▒ļČäĒĢśņŚ¼ ĒĢ£ņ¬ĮņØä ņĀ£Ļ▒░ĒĢ£ Ēøä TSAļź╝ ļČäņŻ╝ĒĢśņśĆļŗż. ĻĘĖļ”¼Ļ│Ā ĻĘĀņŻ╝ļź╝ TSAļ®┤ņŚÉ ĒÜŹņäĀņĀæņóģĒĢśņŚ¼ 28┬░CņŚÉ 10ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņ▓ŁņāēņŚÉņä£ ĒÖ®ņāēņ£╝ļĪ£ ļ│ĆņāēļÉ£ ļČĆļČäņØś Ļ░ĆļĪ£ ņ¦üĻ▓ĮņØä ņĖĪņĀĢĒĢśņśĆļŗż. ļģ╣ļ¦É(1.0%, w/v), ņä¼ņ£Āņåī(CM-cellulose, 0.1% w/v), ņ╣┤ņäĖņØĖ(skimmed milk, 10% w/v), ņ¦Ćņ¦ł(tributyrin, 0.5% v/v)ņØś Ļ░ĆņłśļČäĒĢ┤ļŖźņØĆ SmibertņÖĆ Krieg (1994)ņØś ļ░®ļ▓ĢņŚÉ ņżĆĒĢśņŚ¼ ĒÅēĒīÉņŚÉ ĻĘĀņŻ╝ļź╝ ĒÜŹņäĀ ņĀæņóģĒĢśņŚ¼ 28┬░CņŚÉ 1ņŻ╝ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ņāØņä▒ļÉ£ Ēł¼ļ¬ģĒÖśņØś ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż.
Ļ▓░Ļ│╝
ĒåĀļ¦łĒåĀ ņāØņ£Ī ņ┤ēņ¦ä ĻĘĀņŻ╝ ņäĀļ░£
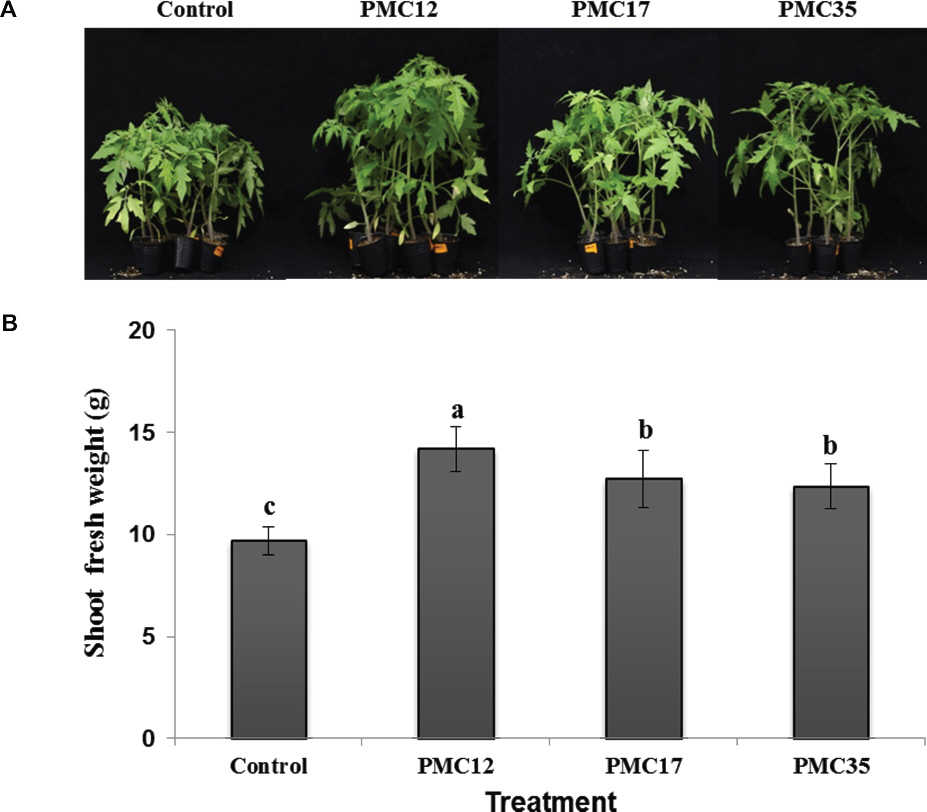
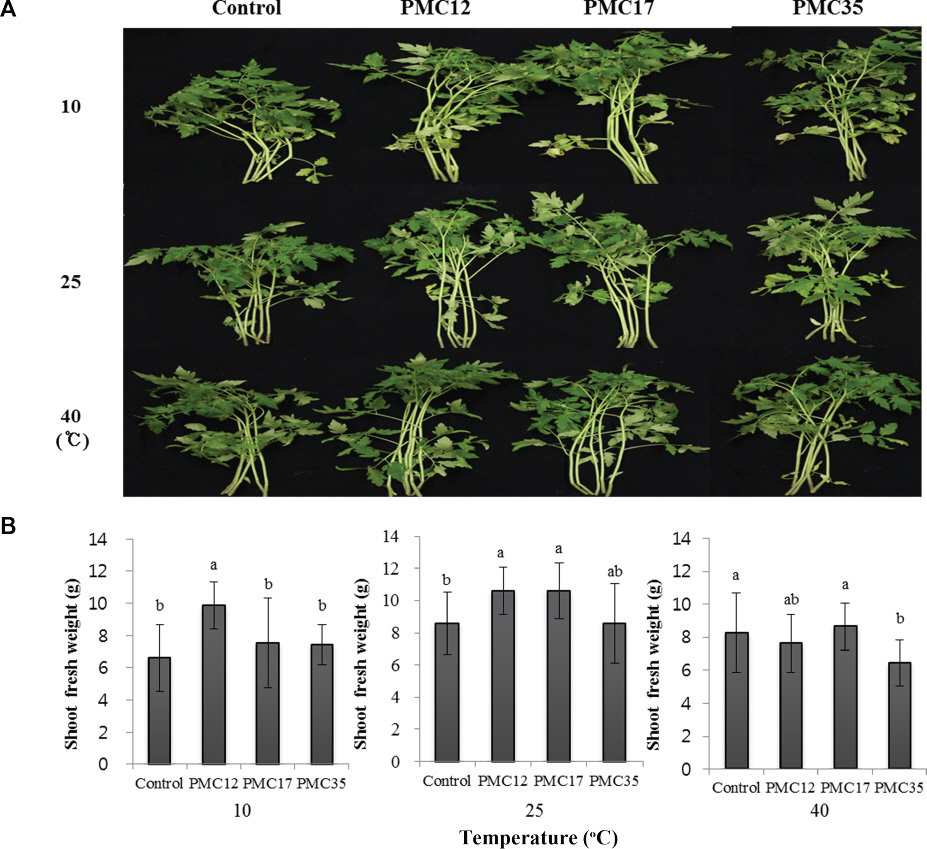
ĒåĀļ¦łĒåĀ ņóģņ×É ļ░Å ĻĘ╝ĻČīņŚÉņä£ 56 ĻĘĀņŻ╝ļź╝ ļČäļ”¼ĒĢśņśĆĻ│Ā ņāØņ£Īņ┤ēņ¦ä Ļ▓ĆņĀĢņØä ĒåĄĒĢ┤ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ĒåĀļ¦łĒåĀņØś ņāØņ▓┤ņżæņØ┤ ĒåĄĻ│äņĀüņ£╝ļĪ£ ņ£ĀņØśņĀüņ£╝ļĪ£ ņ”ØĻ░ĆĒĢ£ PMC12, PMC17, PMC35 ĻĘĀņŻ╝ļź╝ ņäĀļ░£ĒĢśņśĆļŗż(p<0.05). ĒåĀļ¦łĒåĀ ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØĆ PMC12 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ 14.2 gņ£╝ļĪ£ ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļåÆņĢśĻ│Ā, PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ņØś Ļ▓ĮņÜ░ļŖö 12.7 g, 12.4 gņ£╝ļĪ£ ļīĆņĪ░ĻĄ¼ 9.7 g ļ│┤ļŗż ņ£ĀņØśĒĢ£ ņ░©ņØ┤Ļ░Ć ņ׳ņŚłļŗż(Fig. 1).
Fig.┬Ā1
Growth promotion of tomato plants drenched with 10 mM MgSO4 (control), PMC12, PMC17, and PMC35. A representative image (A) and relative shoot fresh weight (B). Experiments were conducted twice with 15 replications. Letters on the bars indicate significant differences by the least significant difference (LSD) test at p<0.05.
ĻĘĀņŻ╝ņØś Ļ│äĒåĄļČäļźś
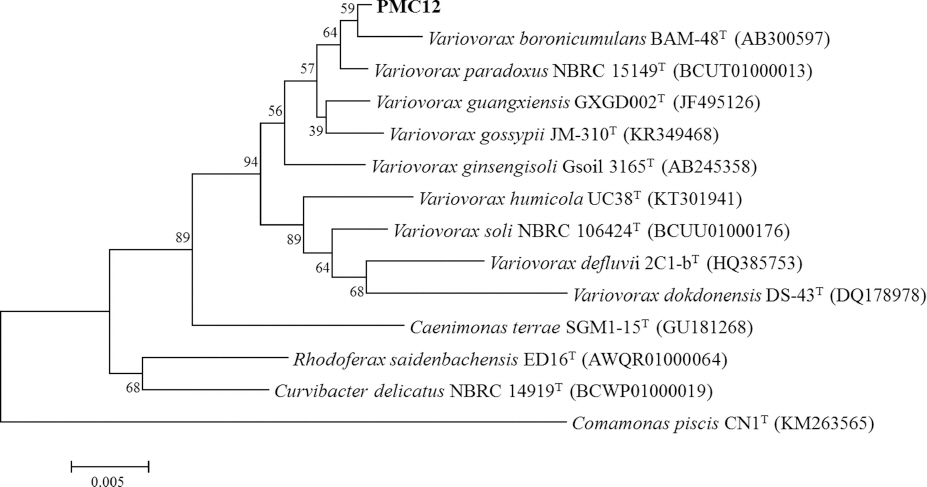
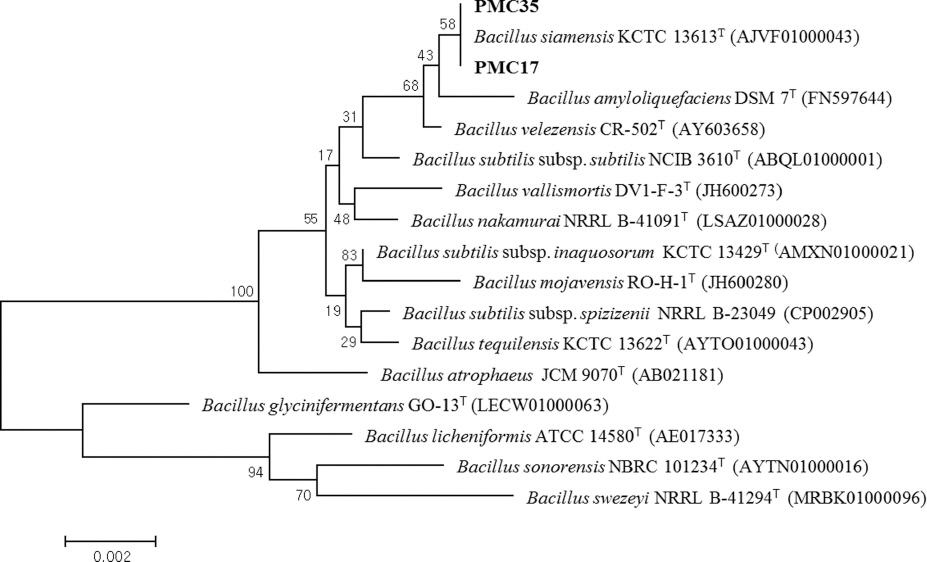
ĒåĀļ¦łĒåĀ ņāØņ£Ī ņ┤ēņ¦ä Ļ▓ĆņĀĢņ£╝ļĪ£ ņäĀļ░£ĒĢ£ PMC12, PMC17, PMC35 ĻĘĀņŻ╝ņØś 16S rRNA gene sequenceļź╝ ĻĖ░ļ░śņ£╝ļĪ£ Ļ│äĒåĄļČäļźśļź╝ ĒĢśņśĆļŗż. PMC12 ĻĘĀņŻ╝ļŖö Variovorax boronicumulans BAM-48T (AB300597)ņÖĆ ņ£Āņé¼ļÅäĻ░Ć 99.66%ļĪ£ ņ£ĀņŚ░Ļ┤ĆĻ│äĻ░Ć Ļ░Ćņן ļåÆņĢśļŗż(Fig. 2). PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ļŖö Bacillus siamensis KCTC 13613T (AJVF01000043)ņÖĆ 16S rRNA ņ£ĀņĀäņ×É ņŚ╝ĻĖ░ņä£ņŚ┤ ņ£Āņé¼ļÅäĻ░Ć 99.93%ļĪ£ ņ£ĀņŚ░Ļ┤ĆĻ│äĻ░Ć Ļ░Ćņן ļåÆņĢśļŗż(Fig. 3).
ĻĘĀņŻ╝ņØś ņāØļ”¼ņāØĒÖöĒĢÖ, ĒśĢĒā£ĒĢÖņĀü ļ░Å ĻĖ░ļŖźņĀü ĒŖ╣ņä▒ ļČäņäØ
PMC ĻĘĀņŻ╝ļŖö R2A, TSA, NA, PDA ļ░░ņ¦ĆņŚÉņä£ ļ¬©ļæÉ ņāØņ£ĪĒĢĀ ņłś ņ׳ņŚłļŗż. ņāØņ£Īņś©ļÅä ļ▓öņ£äļź╝ ņĪ░ņé¼ĒĢ£ Ļ▓░Ļ│╝, PMC12 ĻĘĀņŻ╝ļŖö 2-40┬░C, PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ļŖö 15-50┬░CņŚÉņä£ ņāØņ£ĪņØ┤ Ļ┤Ćņ░░ļÉśņŚłļŗż(Table 1). NaCl ļ▓öņ£äļŖö PMC12 ĻĘĀņŻ╝ļŖö 3%Ļ╣īņ¦Ć, PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ļŖö 10%Ļ╣īņ¦ĆņØś ļåŹļÅäņŚÉņä£ ņāØņ£ĪĒĢĀ ņłś ņ׳ņŚłļŗż. PMC12 ĻĘĀņŻ╝ņØś ņĮ£ļĪ£ļŗł Ēü¼ĻĖ░ļŖö 0.63 mm, PMC17 ĻĘĀņŻ╝ļŖö 1.42 mm, PMC35 ĻĘĀņŻ╝ļŖö 3.33 mmņśĆņ£╝ļ®░ PMC12 ĻĘĀņŻ╝ņØś ņĮ£ļĪ£ļŗł ņāēņØĆ ļģĖļ×Ćņāē, PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ļŖö ĒØ░ņāēņØ┤ņŚłļŗż. Ļ┤æĒĢÖĒśäļ»ĖĻ▓Įņ£╝ļĪ£ Ļ┤Ćņ░░ĒĢ£ PMC12 ĻĘĀņŻ╝ņØś Ēü¼ĻĖ░ļŖö ĒÅŁ 0.5-0.6 ╬╝m, ĻĖĖņØ┤ 2.05 ╬╝m, PMC17 ĻĘĀņŻ╝ļŖö ĒÅŁ 0.7-0.8 ╬╝m, ĻĖĖņØ┤ 3.48 ╬╝mņØ┤ņŚłĻ│Ā PMC35 ĻĘĀņŻ╝ļŖö ĒÅŁ 0.7-0.9 ╬╝m, ĻĖĖņØ┤ 4.6 ╬╝mņØ┤ņŚłļŗż(Table 1). ļģ╣ļ¦É, ņ╣┤ņĀ£ņØĖ, ņä¼ņ£Āņåī, ņ¦Ćņ¦ł, ņØĖņé░ ļō▒ņØś Ļ░ĆņłśļČäĒĢ┤ ņŗżĒŚśņŚÉņä£ PMC12 ĻĘĀņŻ╝ļŖö ĒÖ£ņä▒ņØ┤ ņŚåņŚłĻ│Ā PMC17 ĻĘĀņŻ╝ņÖĆ PMC35 ĻĘĀņŻ╝ļŖö ņ╣┤ņäĖņØĖĻ│╝ ņ¦Ćņ¦ł Ļ░ĆņłśļČäĒĢ┤ļŖźņØ┤ ņ׳ņŚłļŗż. ņŗØļ¼╝ņāØņ£Īņ┤ēņ¦ä Ļ┤ĆļĀ© ĒŖ╣ņä▒ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, ņäĖ ĻĘĀņŻ╝ ļ¬©ļæÉ IAAņāØņä▒ļŖźĻ│╝ ACC ĒāłņĢäļ»╝ĒÜ©ņåī ņāØņä▒ļŖź, ņ¦łņåīĻ│ĀņĀĢ ļ░Å ņĢöļ¬©ļŗłņĢä ņāØņä▒ļŖź, ņŗ£ĒåĀĒéżļŗī ņāØņä▒ļŖźĻ│╝ ņŗ£ļō£ļĪ£ĒżņĢä ņāØņä▒ļŖźņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ ļéśĒāĆļé¼ļŗż. ņŗ£ļō£ļĪ£ĒżņĢä ņāØņä▒ļŖźņØĆ PMC17 ĻĘĀņŻ╝(13 mm)Ļ░Ć Ļ░Ćņן ļåÆņĢśĻ│Ā ļŗżņØīņ£╝ļĪ£ PMC35 ĻĘĀņŻ╝(5.67 mm), PMC12 ĻĘĀņŻ╝(3.67 mm) ņł£ņ£╝ļĪ£ ļé«ņĢśļŗż(Table 1).
Table┬Ā1
Physiological characteristics of strain PMC12, PMC17 and PMC35
ļ╣äņāØļ¼╝ ļ░Å ņāØļ¼╝ ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ņ”ØĻ░Ģ ĒÜ©Ļ│╝
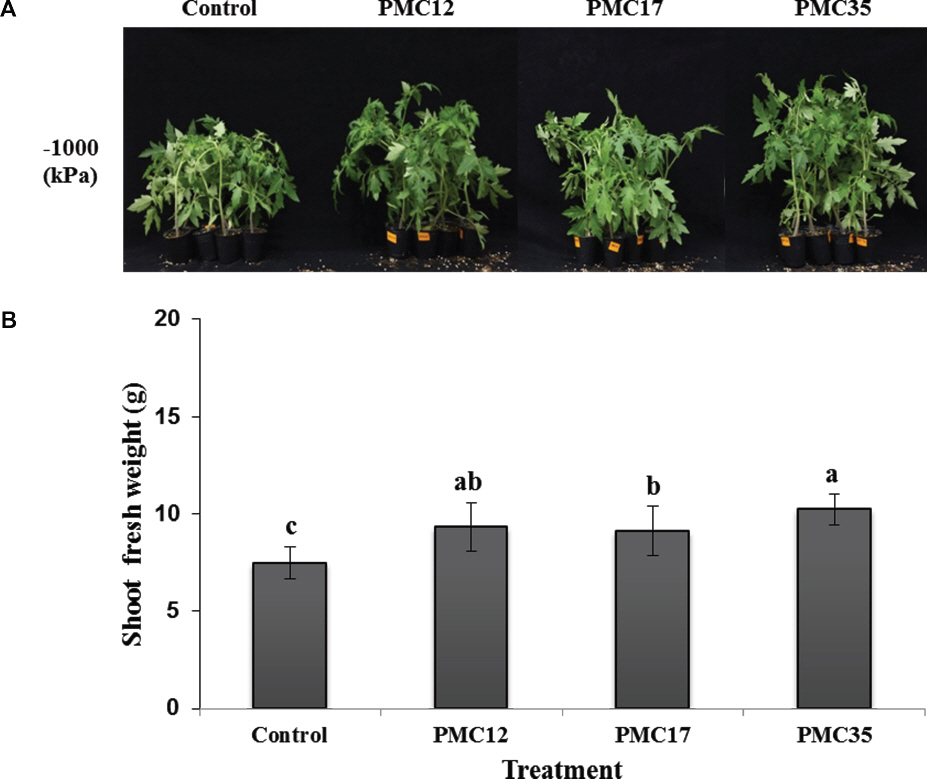
ņŚ╝ņ▓śļ”¼ 2ņŻ╝ Ēøä ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØä ņĖĪņĀĢĒĢ£ Ļ▓░Ļ│╝, ņäĖĻĘĀĒśäĒāüņĢĪņØä Ļ┤ĆņŻ╝ ņ▓śļ”¼ĒĢ£ ņŗżĒŚśĻĄ¼ņŚÉņä£ ļ¬©ļæÉ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņāØņ▓┤ņżæņØ┤ ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļåÆņĢśļŗż(p<0.05). ĒŖ╣Ē׳ PMC35 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ĒåĀļ¦łĒåĀņØś ņāØņ▓┤ņżæņØĆ 10.2 gņ£╝ļĪ£ Ļ░Ćņן ļåÆņĢśĻ│Ā, PMC12 ņ▓śļ”¼ĻĄ¼ļŖö 9.3 g, PMC17 ņ▓śļ”¼ĻĄ¼ļŖö 9.1 gņ£╝ļĪ£ ļīĆņĪ░ĻĄ¼ņØĖ 7.4 g ļ│┤ļŗż ļåÆņĢśļŗż. ļö░ļØ╝ņä£ PMC ĻĘĀņŻ╝ļź╝ ņ▓śļ”¼ĒĢśļ®┤ ņŚ╝ ņŖżĒŖĖļĀłņŖż ņĪ░Ļ▒┤ņŚÉņä£ ņŗØļ¼╝ņ▓┤ņØś ņāØņ£ĪņØ┤ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņ”ØĻ░ĆļÉ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż(Fig. 4).
Fig.┬Ā4
Effect of the strains PMC12, PMC17, and PMC35 on growth of tomato plants treated with -1000 kPa complex salt solution. A representative image (A) and shoot fresh weight (B) of tomato plants after 1 week of salt stress. Letters on the bars indicate significant differences by the least significant difference (LSD) test at p<0.05.
ņś©ļÅä ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ ņ”ØĻ░Ģ ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśĻĖ░ņ£äĒĢ┤ 10, 25, 40┬░CņØś ļ░░ņ¢æĻĖ░ņŚÉ 4ņØ╝Ļ░ä ņś©ļÅä ņŖżĒŖĖļĀłņŖżļź╝ ņ▓śļ”¼ĒĢ£ Ļ▓░Ļ│╝ļŖö Fig. 5Ļ│╝ Ļ░Öļŗż. 10┬░CņØś ņĀĆņś© ņŖżĒŖĖļĀłņŖż ņĪ░Ļ▒┤ņŚÉņä£ PMC12 ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ņāØņ▓┤ņżæņØ┤ 9.8 gņ£╝ļĪ£ ļīĆņĪ░ĻĄ¼ 6.6 gņŚÉ ļ╣äĒĢ┤ ņāØņ▓┤ņżæņØ┤ ņ£ĀņØśņä▒ ņ׳Ļ▓ī ņ”ØĻ░ĆļÉśņŚłļŗż(p<0.05). ĻĘĖļ¤¼ļéś PMC17 ņ▓śļ”¼ĻĄ¼ļŖö 7.5 g, PMC35 ņ▓śļ”¼ĻĄ¼ļŖö 7.4 gņ£╝ļĪ£ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņāØņ▓┤ņżæņØ┤ ņ”ØĻ░ĆļÉśņŚłņ¦Ćļ¦ī ĒåĄĻ│äņĀüņ£╝ļĪ£ ņ£ĀņØśĒĢśņ¦Ć ņĢŖņĢśļŗż. 25┬░C ņĪ░Ļ▒┤ņŚÉņä£ļŖö PMC12 ņ▓śļ”¼ĻĄ¼ņÖĆ PMC17 ņ▓śļ”¼ĻĄ¼ļŖö 10.6 gņ£╝ļĪ£ ņāØņ▓┤ņżæņØ┤ ņ”ØĻ░ĆļÉśņŚłņ¦Ćļ¦ī PMC35 ņ▓śļ”¼ĻĄ¼ļŖö ņ£ĀņØśĒĢ£ ņ░©ņØ┤ļŖö ņŚåņŚłļŗż. 40┬░CņØś Ļ│Āņś© ņŖżĒŖĖļĀłņŖżļź╝ ļ░øņØĆ ĒåĀļ¦łĒåĀļŖö PMC ĻĘĀņŻ╝ļź╝ Ļ┤ĆņŻ╝ņ▓śļ”¼ļź╝ ĒĢśņŚ¼ļÅä ņŖżĒŖĖļĀłņŖżņØś ļé┤ņä▒ņØ┤ ņ”Øņ¦äļÉśņ¦Ć ņĢŖņĢśļŗż. PMC35 ĻĘĀņŻ╝ņØś Ļ▓ĮņÜ░ 6.4 gņ£╝ļĪ£ 8.2 gņØĖ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņāØņ▓┤ņżæņØ┤ ņśżĒ׳ļĀż Ļ░ÉņåīļÉśņŚłļŗż. PMC ĻĘĀņŻ╝ļŖö ņĀĆņś©Ļ│╝ ņżæņś© ņĪ░Ļ▒┤ņŚÉņä£ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņāØņ£ĪņØ┤ ņ”ØĻ░ĆļÉśņŚłņ¦Ćļ¦ī Ļ│Āņś© ņĪ░Ļ▒┤ņŚÉņä£ļŖö ĒåĀļ¦łĒåĀņØś ņāØņ£ĪņØ┤ ņ”ØĻ░ĆĒĢśņ¦Ć ņĢŖņĢśļŗż(Fig. 5B).
Fig.┬Ā5
Effect of the strains PMC12, PMC17, and PMC35 on growth of tomato plants treated at different temperatures. A representative image (A) and shoot fresh weight (B) of tomato plants after 4 days of temperature stress. Letters on the bars indicate significant differences by the least significant difference (LSD) test at p<0.05.
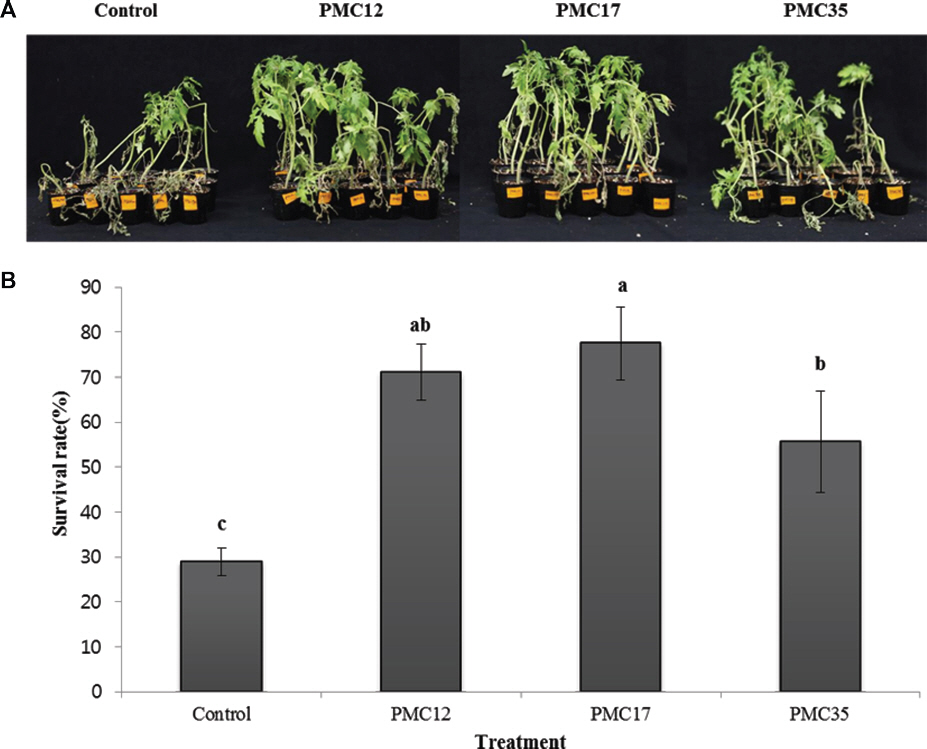
Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ĒåĀļ¦łĒåĀņØś ļé┤ņä▒ ņ”ØĻ░Ģ ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśĻĖ░ ņ£äĒĢ┤ 15ņØ╝Ļ░ä Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖżļź╝ Ļ░ĆĒĢ£ Ļ▓░Ļ│╝ļŖö Fig. 6Ļ│╝ Ļ░Öļŗż. PMC ĻĘĀņŻ╝ļź╝ ņ▓śļ”¼ĒĢśņśĆņØä ļĢī Ļ▒┤ņĪ░ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ļé┤ņä▒ņØ┤ ņ”ØļīĆļÉ©ņØä ĒÖĢņØĖĒĢĀ ņłś ņ׳ņŚłļŗż. ĒåĀļ¦łĒåĀ ņāØņĪ┤ņ£©ņØĆ ļīĆņĪ░ĻĄ¼ 28.9%ņŚÉ ļ╣äĒĢ┤ PMC12 ņ▓śļ”¼ĻĄ¼ļŖö 71.1%, PMC17 ņ▓śļ”¼ĻĄ¼ļŖö 77.6%, PMC35 ņ▓śļ”¼ĻĄ¼ļŖö 55.6%ļĪ£ ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļåÆņĢśļŗż(Fig. 6B).
Fig.┬Ā6
Effect of the strains PMC12, PMC17, and PMC35 on the survival rate of tomato plants after 15 days of drought stress. A representative image (A) and survival rate (B) Experiments were conducted twice with 15 replications. Letters on the bars indicate significant differences by the least significant difference (LSD) test at p<0.05.
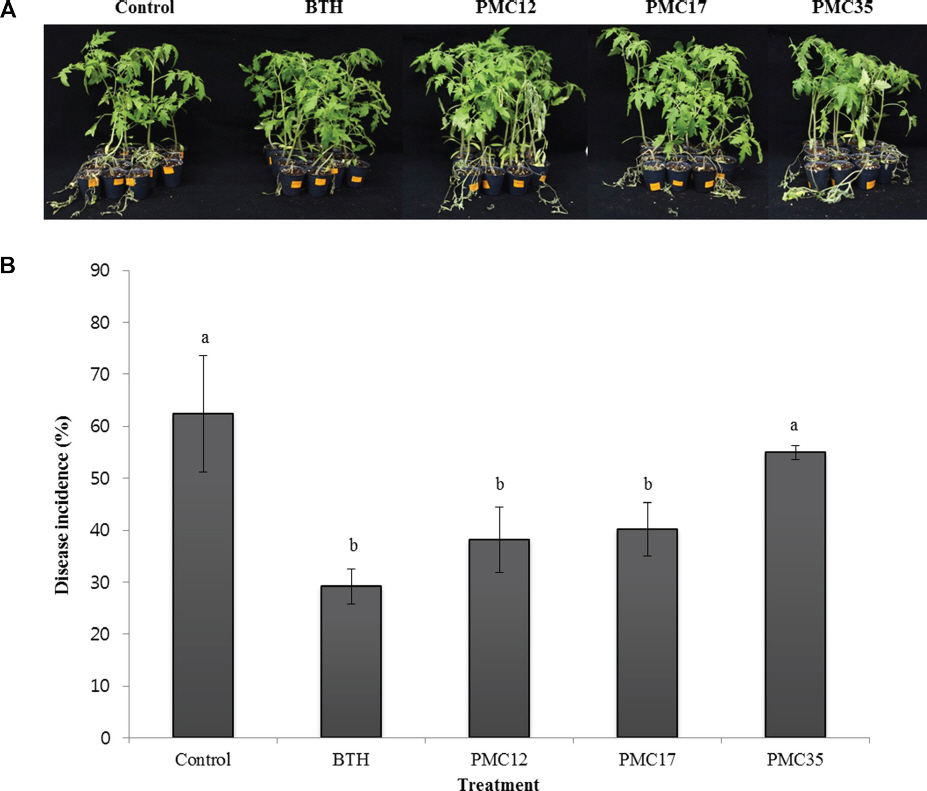
ĒåĀļ¦łĒåĀ ĒÆŗļ¦łļ”äļ│æ ņ¢ĄņĀ£ ĒÜ©Ļ│╝ļŖö ļ│æ ļ░£ņāØņ£©ņØ┤ BTH ņ▓śļ”¼ĻĄ¼ņŚÉņä£ 29.2%ļĪ£ Ļ░Ćņן ļé«ņĢśĻ│Ā PMC12 ĻĘĀņŻ╝ņÖĆ PMC17 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ļŖö Ļ░üĻ░ü 38.1%Ļ│╝ 40.2%ļĪ£ ļīĆņĪ░ĻĄ¼ 62.4% ļ│┤ļŗż ņ£ĀņØśņä▒ ņ׳Ļ▓ī ļé«ņĢśļŗż(p<0.05). ĻĘĖļ¤¼ļéś PMC35 ņ▓śļ”¼ĻĄ¼ņØś ļ│æ ļ░£ņāØņ£©ņØĆ 54.9%ļĪ£ ļīĆņĪ░ĻĄ¼ņÖĆ ņ£ĀņØśĒĢ£ ņ░©ņØ┤ļź╝ ļ│┤ņØ┤ņ¦Ć ņĢŖņĢśļŗż(Fig. 7).
Fig.┬Ā7
Effect of the strains PMC12, PMC17, and PMC35 on the survival of tomato plants treated with Ralstonia solanacearum. A representative image (A) and disease incidence (B) Experiments were conducted twice with 15 replications. Letters on the bars indicate significant differences by the least significant difference (LSD) test at p<0.05.
Ļ│Āņ░░
ļ│Ė ņŚ░ĻĄ¼ļŖö ņŗØļ¼╝ņØś ņāØņ£Ī ņ┤ēņ¦äļ┐Éļ¦ī ņĢäļŗłļØ╝ ļŗżņ¢æĒĢ£ ņāØļ¼╝ ļ░Å ļ╣äņāØļ¼╝ ņŖżĒŖĖļĀłņŖż ļé┤ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżļŖö ĻĖ░ļŖźņØä Ļ░Ćņ¦ä ĻĘĀņŻ╝ļź╝ ļ░£ĻĄ┤ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ĒåĀļ¦łĒåĀ ĻĘ╝ĻČīņ£╝ļĪ£ļČĆĒä░ ĻĘĀņŻ╝ļź╝ ļČäļ”¼ĒĢśņśĆļŗż. ļČäļ”¼ĒĢ£ ĻĘĀņŻ╝ļź╝ ĒåĀļ¦łĒåĀņŚÉ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢ£ Ļ▓░Ļ│╝ PMC12, PMC17, PMC35 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼ņŚÉņä£ ĒåĀļ¦łĒåĀ ņāØņ£ĪņØ┤ ņ”ØļīĆļÉśņŚłļŗż.
PMC12, PMC17, PMC35 ĻĘĀņŻ╝ņØś ņŗØļ¼╝ ņāØņ£Ī ņ┤ēņ¦ä Ļ┤ĆļĀ© ĒŖ╣ņä▒ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, 3 ĻĘĀņŻ╝ ļ¬©ļæÉ ņ¦łņåīĻ│ĀņĀĢļŖź, ņĢöļ¬©ļŗłņĢä ņāØņä▒ļŖź, ACC ĒāłņĢäļ»╝ĒÜ©ņåī ĒÖ£ņä▒, ņŗ£ļō£ļĪ£ĒżņĢä ņāØņä▒ļŖź, ņØĖņé░Ļ░ĆņÜ®ĒÖöļŖź, IAA ļ░Å ņŗ£ĒåĀĒéżļŗī ņāØņä▒ļŖźņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ņŚłļŗż(Table 1). ņŖżĒŖĖļĀłņŖż ņĪ░Ļ▒┤ĒĢśņŚÉņä£ ņŗØļ¼╝ Ļ▓ĆņĀĢĒĢ£ Ļ▓░Ļ│╝ PMC12 ĻĘĀņŻ╝Ļ░Ć ņĀäņ▓┤ņĀüņ£╝ļĪ£ ņÜ░ņłśĒ¢łļŗż.
Variovorax ņåŹņØĆ ļé£ļČäĒĢ┤ņä▒ ņ£ĀĻĖ░ ĒÖöĒĢ®ļ¼╝ņØ┤ļéś ņżæĻĖłņåŹņ£╝ļĪ£ ņśżņŚ╝ļÉ£ ĒÖśĻ▓ĮņŚÉņä£ ĒÆŹļČĆĒĢĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņŗØļ¼╝Ļ│╝ ņ╣£ļ░ĆļÅäĻ░Ć ļåÆņĢä ĻĘ╝ĻČī ļ░Å ļé┤ņāØņäĖĻĘĀņ£╝ļĪ£ļÅä ņĪ┤ņ×¼ĒĢ£ļŗż. ĻĘĖ ņżæ V. paradoxusļŖö ņŗØļ¼╝ ĻĘ╝ĻČīņŚÉņä£ ĒØöĒ׳ ļ░£Ļ▓¼ļÉśļŖö ņל ņĢīļĀżņ¦ä PGPRļĪ£ ņŗØļ¼╝ņØś ņŖżĒŖĖļĀłņŖżļź╝ ņżäņØ┤Ļ│Ā, ņ¢æļČäņØä Ļ░ĆņÜ®ĒÖöņŗ£Ēéżļ®░, ņŗØļ¼╝ ļ│æņøÉĻĘĀņØś ņä▒ņןņØä ņ¢ĄņĀ£ĒĢśļŖö ĻĖ░ļŖźņØ┤ ņ׳ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłļŗż(Han ļō▒, 2013).
ņäĀļ░£ĒĢ£ ĻĘĀņŻ╝ļź╝ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśņśĆņØä ļĢī, ĒåĀļ¦łĒåĀņØś ņāØņ▓┤ņżæņØ┤ ņ”ØĻ░ĆĒĢ£ Ļ▓āņØĆ ĻĘĀņŻ╝Ļ░Ć ņāØņä▒ĒĢ£ ņĢöļ¬©ļŗłņøĆ ņØ┤ņś©ņØä ĒåĀļ¦łĒåĀĻ░Ć ņ¦łņåīņøÉņ£╝ļĪ£ ņé¼ņÜ®Ē¢łĻĖ░ ļĢīļ¼ĖņØĖ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż. Horchani ļō▒ (2010)ņØĆ ĒåĀļ¦łĒåĀĻ░Ć ņĢöļ¬©ļŗłņĢäļź╝ ĒØĪņłśĒĢśņŚ¼ ņāØņ£ĪņØ┤ ņ”ØĻ░ĆļÉ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż. ņØĖņØĆ ņ¦łņåī, ņ╣╝ļź©Ļ│╝ ļŹöļČłņ¢┤ ļ╣äļŻīņØś 3ļīĆ ņÜöņåīļĪ£ ņŗØļ¼╝ņØ┤ ļŗżļ¤ēņ£╝ļĪ£ ĒĢäņÜöļĪ£ ĒĢśņ¦Ćļ¦ī ļīĆļČĆļČä ņØ┤ņÜ®ĒĢśĻĖ░ ņ¢┤ļĀżņÜ┤ ļČłņÜ®ņä▒ņ£╝ļĪ£ ĒåĀņ¢æņŚÉ ņĪ┤ņ×¼ĒĢ£ļŗż. ņØĖņé░ Ļ░ĆņÜ®ĒÖöĻĘĀņØĆ ņ£ĀĻĖ░ņé░ ļō▒ Ēé¼ļĀłņØ┤ĒŖĖ ļ¼╝ņ¦łņØä ļČäļ╣äĒĢśņŚ¼ ļé£ņÜ®ņä▒ ņØĖņé░ņŚ╝ņØä ļČäĒĢ┤ĒĢśļéś ĒåĀņ¢æņŚÉļŖö ņØ╝ļ░śņĀüņ£╝ļĪ£ ĻĘ╝ĻČīņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļŗżļźĖ ļ»ĖņāØļ¼╝Ļ│╝ Ļ▓Įņ¤üĒĢśĻĖ░ņŚÉļŖö ļ░ĆļÅäĻ░Ć ņČ®ļČäĒĢśņ¦Ć ļ¬╗ĒĢ£ Ļ▓ĮņÜ░Ļ░Ć ņ׳ļŗż. Ēśäņ×¼Ļ╣īņ¦Ć ņØĖņé░Ļ░ĆņÜ®ĒÖöļŖźņØ┤ ņ׳ļŖö ņ¦äĻĘĀĻ│╝ ņäĖĻĘĀņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņ×æļ¼╝ņØś ņāØņé░ļ¤ēņØ┤ ņ”ØĻ░ĆĒ¢łļŗżļŖö ņŚ░ĻĄ¼Ļ░Ć ļ│┤Ļ│ĀļÉśņŚłļŗż(De Freitas ļō▒, 1997; ViveganandanņÖĆ Jauhri, 2002). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņ¦łņåīĻ│ĀņĀĢļŖź, ņĢöļ¬©ļŗłņĢä ņāØņä▒ļŖź, ņØĖņé░Ļ░ĆņÜ®ĒÖöļŖźņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ļŖö PMC ĻĘĀņŻ╝ ņ▓śļ”¼ ĒåĀļ¦łĒåĀļŖö ļ╣äļŻīņØś ĒØĪņłśĻ░Ć ļ╣äĻĄÉņĀü ņÜ®ņØ┤Ē¢łĻĖ░ ļĢīļ¼ĖņŚÉ ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņāØņ£ĪņØ┤ ņ”ØĻ░ĆļÉ£ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż.
ņŚ╝, ņĀĆņś©, Ļ▒┤ņĪ░ ļō▒ņØś ĒÖśĻ▓Įļé┤ņä▒ņØä ņ”ØĻ░Ćņŗ£Ēé© ņØ┤ņ£ĀļŖö Ēü¼Ļ▓ī ļæÉ Ļ░Ćņ¦ĆļĪ£ ļ│╝ ņłś ņ׳ļŗż. ļ©╝ņĀĆ PMC ĻĘĀņŻ╝ņØś Ļ┤ĆņŻ╝ņ▓śļ”¼ļĪ£ ņāØņ£ĪņØ┤ ņ┤ēņ¦äļÉśņ¢┤ ļīĆņĪ░ĻĄ¼ļ│┤ļŗż ņŚ¼ļ¤¼ ņŖżĒŖĖļĀłņŖżņŚÉ ļ╣äĻĄÉņĀü ļé┤ņä▒ņØä ņ¦ĆļŗłĻ▓ī ļÉśņŚłņØä Ļ▓āņØ┤ļØ╝ļŖö Ļ▓āĻ│╝ PMC ĻĘĀņŻ╝Ļ░Ć ņāØņä▒ĒĢ£ ņŗØļ¼╝ĒśĖļź┤ļ¬¼ņ£╝ļĪ£ ņØĖĒĢśņŚ¼ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżĻ░Ć Ļ░ÉņåīļÉśņŚłņØä Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż. ņŗØļ¼╝ņØĆ ņ▓┤ļé┤ņŚÉņä£ ņāØņ£ĪņŚÉ ĒĢäņÜöĒĢ£ ņŚ¼ļ¤¼ Ļ░Ćņ¦Ć ĒśĖļź┤ļ¬¼ņØä ĒĢ®ņä▒ĒĢśĻ│Ā ļ»ĖņāØļ¼╝ņŚÉĻ▓īņä£ Ļ│ĄĻĖēļÉśļŖö ņŚ¼ļ¤¼ ĒśĖļź┤ļ¬¼ņŚÉ ļīĆĒĢ┤ņä£ļÅä ņāØļ”¼ņĀüņØĖ ņśüĒ¢źņØä ļ░øļŖöļŗż(Kang ļō▒, 2014). ņśłļź╝ ļōżņ¢┤, ņŗ£ĒåĀĒéżļŗīņØä ņāØņä▒ĒĢśļŖö B. subtilisļŖö ņĖĪļ░▒ļéśļ¼┤ ļé┤ ABA (abscisic acid)ņØś ņ¢æņØä ņĪ░ņĀłĒĢśņŚ¼ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ļé┤ņä▒ņØä ņ£ĀļÅäĒĢ£ļŗż(Liu ļō▒, 2013). ņŗØļ¼╝ņØś ņāØņ£Ī ļ░Å ļ┐īļ”¼ņāØņ£ĪņØä ņ£ĀļÅäĒĢśļŖö IAAļŖö ņ£Āņ╣┤(YUCCA) ļŗ©ļ░▒ņ¦łņŚÉ ņØśĒĢ┤ ĒĢ®ņä▒, ņĪ░ņĀłļÉśļŖöļŹ░ ņŗØļ¼╝ņ▓┤ņŚÉņä£ ļ░£Ēśäļ¤ēņØ┤ ņ”ØĻ░ĆļÉśļ®┤ ĒÖśĻ▓ĮņĀü ņŖżĒŖĖļĀłņŖż ļé┤ņä▒ņØä ņ£ĀļÅäĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Vurukonda ļō▒, 2016). IAAļŖö ņŗØļ¼╝ņ▓┤Ļ░Ć ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżļź╝ ļ░øņØä ļĢī ņäĖĒż ļé┤ņØś ĒÖ£ņä▒ņé░ņåīņØś ņ¢æņØä ņĪ░ņĀłĒĢśņŚ¼ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżņØś ļé┤ņä▒ņØä ņ”ØĻ░Ćņŗ£ĒéżĻ│Ā ņŗ£ĒåĀĒéżļŗīņŚÉ ņØśĒĢ┤ņä£ļÅä Ļ▒┤ņĪ░ ļō▒ņØś ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ļé┤ņä▒ņØä ņ”ØĻ░Ćņŗ£ņ╝░ņØä Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż.
ņŖżĒŖĖļĀłņŖż ņĪ░Ļ▒┤ņŚÉņä£ ņŚÉĒŗĖļĀīņØĆ ņŗØļ¼╝ņØś ņżäĻĖ░ ļ░Å ļ┐īļ”¼ ņāØņ£ĪņØä Ļ░Éņåīņŗ£ĒéżļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Pierik ļō▒, 2007). ņØ╝ļČĆ PGPRņØĆ ņŚÉĒŗĖļĀīņØś ņĀäĻĄ¼ņ▓┤ņØĖ ACCļź╝ ļČäĒĢ┤ĒĢśļŖö ĒÜ©ņåīņØĖ ACC ĒāłņĢäļ»╝ĒÜ©ņåīļź╝ ņāØņä▒ĒĢśņŚ¼, ņŗØļ¼╝ņ▓┤ ļé┤ ņŚÉĒŗĖļĀī ļåŹļÅäļź╝ Ļ░Éņåīņŗ£ņ╝£ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁļĀźņØä ļåÆņŚ¼ņżĆļŗż(Glick, 2005). ACC ĒāłņĢäļ»╝ĒÜ©ņåīļź╝ ņāØņä▒ĒĢśļŖö PseudomonasļŖö ņŚ╝ņØ┤ļéś Ļ▒┤ņĪ░ ņĪ░Ļ▒┤ņŚÉņä£ ņ×æļ¼╝ ņŚ┤ļ¦żņØś ņä▒ņłÖņØä ņ┤ēņ¦äņŗ£Ēéżļ®░ ņłśĒÖĢļ¤ēņØä ņ”ØĻ░Ćņŗ£ĒéżĻ│Ā, Bacillus, Variovorax ļō▒ ACC ĒāłņĢäļ»╝ĒÜ©ņåīļź╝ ņāØņä▒ĒĢśļŖö ļŗżņ¢æĒĢ£ PGPRņØ┤ ņŗØļ¼╝ņØś ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖż ņÖäĒÖö ļ░Å ļé┤ņä▒ ņ£ĀļÅäņÖĆ ņŚ░Ļ┤ĆņØ┤ ņ׳ļŗżĻ│Ā ļ│┤Ļ│ĀļÉśņŚłļŗż(Arshad ļō▒, 2008; Saleem ļō▒, 2007). ACC ĒāłņĢäļ»╝ĒÜ©ņåīņŚÉ ņØśĒĢ┤ņä£ ņŚÉĒŗĖļĀī ņāØņä▒ņØ┤ Ļ░ÉņåīļÉśĻ│Ā Ļ▓░Ļ│╝ņĀüņ£╝ļĪ£ ĒåĀļ¦łĒåĀĻ░Ć ļ░øļŖö ņŚ╝, ņś©ļÅä, Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖżĻ░Ć Ļ▓ĮĻ░ÉļÉśņŚłņØä Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņØ┤ņÜ®ĒĢ£ PMC ĻĘĀņŻ╝ļŖö in vitro ņŗżĒŚśņØä ĒåĄĒĢ┤ņä£ ACC ĒāłņĢäļ»╝ĒÜ©ņåī, IAA, ņŗ£ĒåĀĒéżļŗī ĒśĖļź┤ļ¬¼ņØä ņāØņä▒ĒĢ©ņØä ĒÖĢņØĖĒĢśņśĆļŗż.
ļīĆļČĆļČäņØś ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ļŖö ĒĢŁĻĘĀĻĖ░ņ×æņŚÉ ĻĖ░ņ┤łĒĢ£ ļ░®ļ▓ĢņØ┤ņŚłņ£╝ļéś ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ļŖźļĀźņØ┤ ņ׳ļŖö ĻĘ╝ĻČīņäĖĻĘĀ ņżæ ņŗØļ¼╝ņŚÉ ņ£ĀļÅäņĀĆĒĢŁņä▒ņØä ņØ╝ņ£╝ĒéżļŖö ĻĖ░ņ×æņØ┤ ņĢīļĀżņ¦Ćļ®┤ņä£ ņ£ĀļÅäņĀĆĒĢŁņä▒ ļŖźļĀźņØ┤ ņÜ░ņłśĒĢ£ ĻĘ╝ĻČīļ»ĖņāØļ¼╝ņØä ņäĀļ░£ĒĢśņŚ¼ ņŗØļ¼╝ļ│æņØś ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņŚÉ ņØ┤ņÜ®ĒĢśĻĖ░ ņ£äĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ņłśĒ¢ēļÉśņŚłļŗż(Van Loon ļō▒, 1998). Leeman ļō▒ (1995)ņØĆ Pseudomonas fluorescens wc374ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņ×ÉņŚ░ Ļ░ÉņŚ╝ļÉ£ ļ╣äļŗÉĒĢśņÜ░ņŖżļé┤ņØś ļ¼┤ ņŗ£ļōżņØīļ│æņØä 50% ņØ┤ņāü ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ņŚłļŗż. PGPRņØä Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśļ®┤ ĒåĀņ¢æ ļ»ĖņāØļ¼╝ ĻĄ░ņ¦æņØ┤ ļ│ĆĒĢśĻ▒░ļéś ĻĘ╝ĻČī ļō▒ņŚÉ ņÜ░ņĀÉĒĢśņŚ¼ ļ│æņøÉĻĘĀņØś ņ╣©ņ×ģņØä ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ļŗż(Mendes ļō▒, 2013). ņŗ£ļō£ļĪ£ĒżņĢä ņāØņä▒ļŖź ļśÉĒĢ£ ļ│æ ļ░£ņāØņ¢ĄņĀ£ņŚÉ ĻĖ░ņŚ¼ĒĢ£ Ļ▓āņ£╝ļĪ£ ņČöņĖĪļÉ£ļŗż. Pseudomonasļź╝ ņØ┤ņÜ®ĒĢ£ ņØ╝ļČĆ ņŚ░ĻĄ¼ņŚÉņä£ ņ▓ĀļČäņŚÉ ļīĆĒĢ£ ĒÜ©Ļ│╝ņĀüņØĖ Ļ▓Įņ¤üņØ┤ ņ¦łļ│æ ņ¢ĄņĀ£ņŚÉ ĻĖ░ņŚ¼ĒĢĀ ņłś ņ׳ļŗżļŖö Ļ▓░Ļ│╝Ļ░Ć ņ׳ņŚłņ¦Ćļ¦ī, Ba┼¤arņÖĆ Bhat (2004)ņØś Ļ▓░Ļ│╝ņŚÉ ņØśĒĢśļ®┤ ņŗ£ļō£ļĪ£ĒżņĢä ņāØņé░ņØĆ R. solanacearumņŚÉļŖö ņśüĒ¢źņØ┤ ņŚåņŚłļŗż. ņĢäņÜĖļ¤¼ V. paradoxusļŖö quorum sensingĻ│╝ Ļ┤ĆļĀ©ļÉ£ ļ»ĖņāØļ¼╝ ņŗĀĒśĖ ņĀäļŗ¼ ļČäņ×É N-acyl homoserine lactone (AHL)ņØä ļČäĒĢ┤ĒĢĀ ņłś ņ׳ļŗż(LeadbetterņÖĆ Greenberg, 2000)ļŖö Ļ▓░Ļ│╝Ļ░Ć ņ׳ļŖöļŹ░, ņĪ┤ņ×¼ĒĢśļŖö ļ│æņøÉņä▒ ĻĘĀņŻ╝ņØś ļÅģņä▒ņØä Ļ░Éņåīņŗ£ĒéżļŖö quorum quenchingņ£╝ļĪ£ ļ│æņøÉņä▒ Ļ░ÉņŚ╝ņ£╝ļĪ£ļČĆĒä░ ĻĖ░ņŻ╝ ņŗØļ¼╝ņØä ļ│┤ĒśĖĒ¢łļŗżļŖö ļ│┤Ļ│ĀĻ░Ć ņ׳ļŗż(Chen ļō▒, 2013). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļ│æņøÉĻĘĀņØä ņĀæņóģĒĢśĻĖ░ 1ņŻ╝ņØ╝ ņĀä PMC ĻĘĀņŻ╝ļź╝ Ļ┤ĆņŻ╝ņ▓śļ”¼ ĒĢśņśĆļŖöļŹ░, PMC ĻĘĀņŻ╝ņŚÉ ņØśĒĢ┤ ĒÆŗļ¦łļ”äļ│æĻĘĀņØä ņ¢ĄņĀ£ĒĢśļŖö ĒÜ©Ļ│╝ļź╝ Ļ░Ćņ¦ä ļ»ĖņāØļ¼╝ ĻĄ░ņ¦æņØ┤ ņ£ĀļÅäļÉśĻ▒░ļéś, ĻĘ╝ĻČīņŚÉ ņÜ░ņĀÉĒĢśņŚ¼ ļ│æņøÉĻĘĀņØ┤ ņŗØļ¼╝ņ▓┤ņŚÉ ņ╣©ņ×ģĒĢśĻĖ░ ņ¢┤ļĀĄĻ▓ī ĒĢśņśĆņØä Ļ░ĆļŖźņä▒ņØ┤ ņ׳ļŗż.
Ēśäņ×¼ ņ¦ĆĻĄ¼ņś©ļé£ĒÖöļĪ£ ņØĖĒĢ£ ĻĖēĻ▓®ĒĢ£ ĒÖśĻ▓Įļ│ĆĒÖöļĪ£ ņŗØļ¼╝ ĒÖśĻ▓Į ņŖżĒŖĖļĀłņŖżņØś Ļ░ĢļÅäņÖĆ ļ│æĒĢ┤ņČ®ņØś ļ░£ņāØļ╣łļÅäĻ░Ć ņĀÉņ░© ļŖśņ¢┤Ļ░ĆĻ│Ā ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņäĀļ░£ļÉ£ PMC12ļŖö ņŗØļ¼╝ņāØņ£ĪĻ│╝ ņŚ╝, ņś©ļÅä, Ļ▒┤ņĪ░ ļō▒ņØś ļ╣äņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖżņÖĆ ĒåĀļ¦łĒåĀ ĒÆŗļ¦łļ”äļ│æ ņ¢ĄņĀ£ ļō▒ ļäōņØĆ ĒÜ©Ļ│╝ ļ▓öņ£äļź╝ Ļ░Ćņ¦äļŗż. PMC12ļŖö ĻĖēļ│ĆĒĢśļŖö ļåŹņŚģĒÖśĻ▓ĮņŚÉņä£ļÅä ņāØĒā£Ļ│äņØś ĻĘĀĒśĢņØä Ļ╣©ĒŖĖļ”¼ņ¦Ć ņĢŖĻ│Ā ņĢłņĀäĒĢ£ ļåŹņé░ļ¼╝ņØä ņāØņé░ĒĢśļŖöļŹ░ ĻĖ░ņŚ¼ĒĢĀ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ĻĖ░ļīĆļÉ£ļŗż.
ņÜöņĢĮ
ĻĘ╝ĻČīņäĖĻĘĀņØĆ ņŗØļ¼╝ ņāØņ£ĪĻ│╝ Ļ▒┤Ļ░Ģ ņ”Øņ¦äņŚÉ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢśļ®░ ņāØļ¼╝ĒĢÖņĀü ņŖżĒŖĖļĀłņŖżļ┐Éļ¦ī ņĢäļŗłļØ╝ ņĀĆņś©, Ļ│Āņś©, Ļ▒┤ņĪ░ ļ░Å ņŚ╝Ļ│╝ Ļ░ÖņØĆ ļ╣äņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖżņŚÉļÅä ļé┤ņä▒ņØä ļČĆņŚ¼ĒĢ£ļŗż. ļ│Ė ņŚ░ĻĄ¼ļŖö ĒåĀļ¦łĒåĀņŚÉ ņāØļ¼╝ņĀü ļ░Å ļ╣äņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖżļź╝ ņÖäĒÖöņŗ£ĒéżļŖö ĻĖ░ļŖźņØä Ļ░Ćņ¦ä ņŗØļ¼╝ ņāØņןņ┤ēņ¦ä ĻĘ╝ĻČīņäĖĻĘĀ(plant growth promoting rhizobacteria, PGPR)ņØä ņäĀļ░£ĒĢśļŖö Ļ▓āņØä ļ¬®Ēæ£ļĪ£ ĒĢśņśĆņ£╝ļ®░ ĒåĀļ¦łĒåĀ ĻĘ╝ĻČīņŚÉņä£ Variovorax sp. PMC12ĻĘĀņŻ╝ļź╝ ļČäļ”¼ĒĢśņśĆļŗż. PMC12ĻĘĀņŻ╝ļŖö in vitroņŚÉņä£ PGPRņØś ĒŖ╣ņä▒ņ£╝ļĪ£ ņĢīļĀżņ¦ä ņĢöļ¬©ļŗłņĢä, IAA, ņŗ£ļō£ļĪ£ĒżņĢä ļ░Å ACC ĒāłņĢäļ»╝ĒÜ©ņåīļź╝ ņāØņä▒ĒĢśņśĆļŗż. PMC12 ĻĘĀņŻ╝ļź╝ ņ▓śļ”¼ĒĢ£ ĒåĀļ¦łĒåĀļŖö ļīĆņĪ░ĻĄ¼ņŚÉ ļ╣äĒĢ┤ ņŚ╝, ņĀĆņś© ļ░Å Ļ▒┤ņĪ░ ņŖżĒŖĖļĀłņŖż ņĪ░Ļ▒┤ņŚÉņä£ ņ¦ĆņāüļČĆ ņāØņ▓┤ņżæņØ┤ ņ£ĀņØśņĀüņ£╝ļĪ£ ļåÆņĢśļŗż. ļśÉĒĢ£ PMC12 ĻĘĀņŻ╝ļź╝ ņ▓śļ”¼ĒĢ£ ĒåĀļ¦łĒåĀļŖö Ralstonia solanacearumņŚÉ ņØśĒĢ£ ņäĖĻĘĀņä▒ ņŗ£ļōżņØīļ│æņŚÉ ļīĆĒĢ£ ņĀĆĒĢŁņä▒ņØ┤ ņ”ØĻ░ĆļÉśņŚłļŗż. Ļ▓░Ļ│╝ņĀüņ£╝ļĪ£ PMC12 ĻĘĀņŻ╝ļŖö ņŗØļ¼╝ņØś ļ╣äņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖż ļ░Å ņāØļ¼╝ņĀü ņŖżĒŖĖļĀłņŖżņŚÉ ļīĆĒĢ£ Ļ░Éņłśņä▒ņØä Ļ░Éņåīņŗ£ĒéżļŖö ņ£Āļ¦ØĒĢ£ ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņĀ£ ļ░Å ņāØļ¼╝ĒÖ£ņä▒ņĀ£ļĪ£ ņé¼ņÜ®ļÉĀ ņłś ņ׳ņØä Ļ▓āņ£╝ļĪ£ ņĀäļ¦ØļÉ£ļŗż.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






