

서론
배추는 김치의 주원료로 전국 각지에서 재배되고 있으며, 엽채류 재배면적 가운데 1순위로 전체 재배면적이 약 25,576 ha에 이른다. 우리나라에서 배추는 봄과 가을로 작기를 구분하고 있으며, 봄배추의 약 54%가 강원도에서 생산되고 있다 (Ministry of Agriculture, Food and Rural Affairs, 2013). 그러나, 배추는 무사마귀병(Plasmodiophora brassicae), 질경이모자이크바이러스(Ribgrass mosaic virus, RMV) 등의 다양한 병해로 인해 경제적으로 큰 피해를 받고 있다(Kim 등, 2003, 2012). 강원도 태백시의 고랭지배추 재배지에서 관리병해충으로 지정된 사탕무씨스트선충(Heterodera schachtii)에 의한 피해가 2011년 최초로 확인되었으며, 그 피해가 인근지역인 삼척시와 정선군으로 확산되고 있다(Kim 등, 2015). 한편 삼척시 고랭지배추 재배지 일부에서 배추에 피해를 일으키지 않는 콩씨스트선충(H. glycines)도 검출되어, 방제 의사결정에 정확한 종 동정을 필요로 하고 있다(Kim 등, 2015).
사탕무씨스트선충(H. schachtii)은 겨자과, 명아주과 등 9과 23속 200여종의 식물이 기주로 알려져 있으며(Steele, 1965), 유럽, 중동, 미국 등 많은 나라에서 문제되고 있는 식물기생선충 가운데 하나이다(Sharma, 1998). 반면, 콩씨스트선충(H. glycines)은 콩과 등 일부 작물에만 피해를 일으키는 제한적인 기주범위를 가진다. 이들은 1980년대부터 1990년대까지 미국 최대 콩 생산지인 오하이오주에서 경제적으로 큰 손실을 일으킨 바 있으며(Willson 등, 1996), 일본, 중국과 브라질 등에서 도 큰 피해를 일으키고 있다(Sharma, 1998). 그러나 2종 모두 Heterodera 속의 ‘Schachtii’ 그룹(Sensu stricto group)에 속하는 씨스트선충으로 클로버씨스트선충(H. trifolii)과 같은 근연종들 간 형태적 특징이 매우 유사하여 종 동정에 많은 어려움을 겪고 있다(Starr 등, 2002; Subbotin 등, 2010).
선충의 방제 의사 결정을 위해서는 정확하고 신속한 종 동정이 필요하다. Powers (2004)와 Subbotin 등(2010)에 따르면, Heterodera 속을 포함한 식물기생선충의 종 동정에는 주로 체장, 체폭, 구침과 생식기 등 각 형태에 대한 길이, 폭 등의 평균 측정치가 이용되어 왔다. 그러나, 형태적 특징을 이용한 종 동정에는 많은 시간, 노력과 노하우를 필요로 하며, 하나의 포장에 2종 이상의 씨스트선충이 혼합되어 있는 경우 종 동정에 많은 어려움이 발생한다(Powers, 2004). 따라서, 최근에는 신속하고 정확한 종 동정을 위해 제한효소절편길이다형성(Restriction fragment length polymorphism with PCR product, PCR-RFLP), SCAR (Sequence characterised amplified region) 및 다중중합효소연쇄반응(Multiplex PCR) 등과 같은 다양한 기법이 진단에 이용되고 있다. PCR-RFLP 기법은 Heterodera 속 씨스트선충의 종 동정에 중요한 분류키로 이용되어 왔으며, ITS rRNA 영역이 주로 이용되어 왔다(Amiri 등, 2002; Maafi 등, 2003; Subbotin 등, 2000). 종 특이 프라이머(Species specific primer)를 이용하면 특정 종의 선충을 신속하게 동정할 수 있다. Heterodera 속 씨스트선충에 대한 종 특이 프라이머는 Amiri 등(2002)에 의해 ITS rRNA 영역을 대상으로 연구되기 시작하였고, 최근에는 18S ribosomal RNA, Actin 1과 mitochondrial DNA의 COI (mtDNA COI) 등 다양한 유전자 영역이 종 특이 프라이머 연구에 이용되고 있다(Toumi 등, 2013a, 2013b).
mtDNA COI 유전자 영역은 해양선충과 동물기생선충의 분류 동정을 위한 DNA 바코드 연구에 이용되고 있으며(Derycke 등, 2010; Prosser 등, 2013), ITS rRNA 영역의 염기서열 비교로 종 구분이 어려웠던 Heterodera 속 ‘Schachtii’ 그룹 5종의 씨스트선충을 뚜렷하게 동정할 수 있었다(Vovlas 등, 2015). 본 연구에서는 Heterodera 속 씨스트선충 mtDNA COI 유전자 영역의 DNA 염기서열 차이를 이용하여 국내 고랭지배추 재배지에서 검출되고 있는 사탕무씨스트선충과 콩씨스트선충을 정확하고 신속하게 동정할 수 있는 기술을 개발하고자 하였다.
재료 및 방법
씨스트선충 채취 및 씨스트 분리
강원도 삼척시 PM001과 BC115의 2개 지역과 정선군 GC147, GC408, YS224와 DA142의 4개 지역 고랭지배추 포장에서 2016년 7월 27일과 9월 13일 총 2회에 걸쳐 씨스트선충이 감염되어 있는 토양시료를 채취하였다(Table 1). 토양시료는 모종삽을 이용하여 100평당 10개 지점 기준으로 깊이 20 cm 이내 작토층의 토양을 채취하였다. 채취한 토양은 골고루 섞은 다음 플라스틱 비커를 이용하여 500 cm3 토양을 정량하고 10 l 물이 들어있는 플라스틱 통에 넣어 토양현탁액을 만들었다. 토양현탁액을 20 mesh, 60 mesh 체에 차례로 거른 다음, 60 mesh 체 위에 남은 물질은 세척통을 이용하여 비커로 옮겨 담았다. 비커에 옮겨 담은 용액을 150 mm 필터페이퍼를 이용하여 거른 다음, 실체현미경 아래서 핀셋으로 갈색의 건전한 씨스트만 골라 유리 고정병에 넣은 후 4°C 냉장고에 보관하여 실험에 사용였다.
Table 1
The legend of the soil samples collected from Samcheok and Jeongseon for identification
씨스트선충의 DNA 추출
씨스트선충의 DNA 추출은 Iwahori 등(2000)의 방법을 이용하였다. 실체현미경 아래서 핀셋으로 씨스트를 골라내어 증류수가 한방울 떨어뜨려져 있는 슬라이드글라스 위에 옮겼다. 골라낸 씨스트를 소형 핀셋을 이용하여 필터페이퍼 조각(2 mm×2 mm 이하)으로 압 력을 가하여 씨스트를 깨뜨렸다. 깨뜨린 씨스트 속에 들어 있는 2기 유충과 알을 필터페이퍼로 압력을 가하여 파쇄시킨 다음, 그 조각을 Proteinase K 용액(2 M KCl2, 10 mM Tris- HCl, 1 M MgCl2, 10% [w/v] Triton-X 100, 20.6 mg/ml Proteinase K) 20 µl가 들어있는 0.2 ml PCR 튜브에 넣었다. PCR cycler (MJ Research, PTC-200, USA)를 이용하여 60°C에서 30 분, 94°C에서 10분간 Lysis 과정을 거쳐 DNA를 추출하였다.
MtDNA COI 유전자의 중합효소연쇄반응
씨스트선충의 mtDNA COI 유전자 영역에 대한 중합효소연쇄반응(PCR)을 수행하였다. mtDNA COI 유전자 영역의 PCR을 위하여 forward primer #JB3 (5’-TTTTTTGGGCATCCTGAGGTTTAT-3’)와 reverse primer #JB5 (5’-AGCACCTAAACTTAAAACATAATGAAAATG-3’) 를 이용하였다(Derycke 등, 2005). PCR 반응은 premix (Nanohelix™, HelixAmp™ Ready-2×-Go, Daejeon, Korea) 15 µl가 들어있는 0.2 ml PCR 튜브에 template DNA 4 µl, 10 pmol의 forward와 reverse primer 각 0.5 µl, Triple distilled water 30 µl를 넣고 혼합한 다음 PCR cycler (PTC-200; MJ Research, Alameda, CA, USA)를 이용하여 수행하였으며, 94°C에서 30초간 denaturation, 53°C에서 30초간 anealing, 72°C에서 45초간 extension의 과정을 40회 반복하였다. PCR 증폭산물은 1% 아가로오스겔과 1×TAE buffer (0.04 M Tris-Acetate, 0.001 M EDTA) 에서 100 V, 25 mA로 25분간 전기영동한 다음 UV장치(UVCI- 1100; Major science, New Taipei City, Taiwan)에서 DNA의 증폭여부를 확인하였다.
씨스트선충의 DNA 염기서열 분석 및 계통수 작성
유전자 증폭산물은 PCR 정제키트(Macherey-Nagel, Gel and PCR Clean Up)를 이용하여 정제한 다음, (주)제노텍 유전체분석센터(Daejeon, Korea)에 DNA 염기서열 분석을 의뢰하였다. 씨스트선충의 mtDNA COI 유전자의 염기서열은 미국국립생물정보 센터(NCBI)에 기등록된 데이터들과 함께 Clustal×1.83 기본값으로 정렬하였으며, Mrbayes 3.2.6의 General Time Reversible substitution model (GTR+G)을 이용하여 분자생물학적 계 통수를 작성하였다. 계통수 작성을 위한 외집단분류군(Outgroup)의 선발은 Vovlas 등(2015)의 연구 결과에 따랐다. 종내 변이율과 종간변이율은 Mega 6.06 프로그램의 Distance 분석을 통해 확인하였다.
PCR 산물의 제한효소절편길이다형성
Mitochondrial DNA COI 유전자의 제한효소 처리에 따른 절편길이다형성(RFLP)을 확인하기 위하여 PCR 증폭산물에 PstI, VspI, AlwI, RsaI, MvaI, EcoRI, Eco72I과 HinfI 8종류의 제한효소를 처리하였다. 제한효소의 반응을 위해 0.2 ml의 PCR 튜브에 mtDNA COI 유전자 증 폭산물 3 µl, Triple distilled water 5.5 µl, Restriction enzyme 0.5 µl (5 unit/µl)과 각 제한효소의 10× buffer 1 µl를 넣고 혼합 한 다음 37°C에서 3시간 동안 배양하였다. 제한효소 반응액은 3% agarose gel, 1× TAE buffer (0.04 M Tris-Acetate, 0.001 M EDTA)에서 100 V, 25 mA로 40분간 전기영동한 다음, UV장치 (UVCI-1100; Major science)에서 DNA 밴드양상을 확인하였다 (Kim 등, 2014).
씨스트선충 2종의 특이 프라이머 제작
종 특이 프라이머 제작은 Seo 등(2012)의 방법을 이용하였다. NCBI에 기등록되어 있는 사탕무씨스트선충(H. schachtii)과 콩씨스트선충(H. glycines) mtDNA COI 유전자 영역의 448 bp 염기서열을 H. trifolii (KT163239), H. ciceri (KC172919), H. elachista (KC618472), H. daverti (KT163237)의 mtDNA COI 유전자 영역 염기서열과 함께 Mega 6.06 프로그램의 기본값(default)으로 정렬하였다. 정렬한 결과를 비교하여 사탕무씨스트선충 또는 콩씨 스트선충에만 특이적으로 존재하고 다른 종에는 없는 뉴클레오티드를 3’ 말단 염기로 하여, 사탕무씨스트선충만 특이적으로 작동하는 forward primer #JBS1 (5’-GGATAATTTATGCTATTATC- 3’)과 콩씨스트선충만 특이적으로 작동하는 forward primer #JBG1 (5’-TGGTTTAGTTAGATTAACTATC-3’)을 설계하였다. Reverse primer는 염기서열 비교에 이용한 모든 종에 동일하게 존재하는 염기서열을 이용하여 #JB3R (5’-TCCAAACTWGCGTTACTYAG- 3’)을 설계하였다. PCR 프라이머의 합성은 (주)제노텍(Daejeon, Korea)에 의뢰하였다.
종 특이 프라이머를 이용한 PCR
사탕무씨스트선충(H. schachtii)과 콩씨스트선충(H. glycines)의 종 판별을 위해 본 연구에서 설계된 PCR 프라이머 세트(#JBS1, #JBG1, #JB3R)를 이용하여 씨스트(cyst)로부터 추출된 DNA를 주형으로 PCR을 수행 하였다. 해당 프라이머 세트가 사탕무씨스트선충과 콩씨스트선충에만 특이적으로 반응하는지 여부를 확인하기 위해, 국립 농업과학원 작물보호과에서 유지증식 중인 고구마뿌리혹선충 (Meloidogyne hapla), 땅콩뿌리혹선충(M. arenaria), 당근뿌리 혹선충(M. hapla), 딸기뿌리썩이선충(Pratylenchus penetrans), 사과뿌리썩이선충(P. vulnus)과 3차 증류수(Negative control)도 함께 PCR 반응시켰다. PCR 반응은 premix (Nanohelix™, HelixAmp™ Ready-2×-Go) 15 µl가 들어있는 0.2 ml PCR 튜브에 template 4 µl, 10 pmol의 3종류 프라이머(#JBS1, #JBG1, #JB3R)를 각각 1 µl씩 총 3 µl, 3차 증류수 28 µl를 넣고 혼합한 다음 PCR cycler (PTC-200; MJ Research)를 이용하여 수행하였으며, 94°C에서 30초간 denaturation, 44°C에서 30초간 anealing, 72°C에서 45초간 extension의 과정을 40회 반복하였다. PCR 증폭산물은 2.5% 아가로오스겔과 1×TAE buffer (0.04 M Tris-Acetate, 0.001 M EDTA)에서 100 V로 25분간 전기영동한다음, UV장치(UVCI-1100; Major science)에서 DNA의 증폭여부를 확인하였다.
DNA 농도에 따른 종 특이 프라이머의 PCR 민감도
사탕무 씨스트선충(H. schachtii, GC147)과 콩씨스트선충(H. glycines, YS224)의 DNA 농도에 따른 종 특이 프라이머의 PCR 민감도를 검정하기 위해, 씨스트로부터 추출한 DNA는 Choi 등(2014)의 방법을 응용하여 10배씩 연속 희석하여 1 µl 당 100 ng, 10 ng, 1 ng, 100 pg, 10 pg과 1 pg으로 조정하였다. PCR 반응은 premix (Nanohelix™, HelixAmp™ Ready-2×-Go) 15 µl가 들어있는 0.2 ml PCR 튜브에 template DNA 1 µl, 10 pmol의 3종류 프라이머(#JBS1, #JBG1, #JB3R)를 각각 1 µl씩 총 3 µl, Triple distilled water 31 µl를 넣고 혼합한 다음 PCR cycler (PTC-200; MJ Research)를 이용하여 수행하였다.
결과 및 고찰
씨스트선충 mtDNA COI 유전자 영역의 PCR 결과
강원도 삼척 2개와 정선 4개의 고랭지배추 재배지에서 검출된 씨스트선충의 종 동정을 위해 mtDNA COI 유전자의 PCR을 수행하였다. 이를 위해 총 6개의 포장에서 씨스트를 각각 10개씩 표본집단으로 건져 DNA를 추출하고 PCR을 수행하였다. 그 결과 6개 포장에서 검출된 씨스트선충에서 모두 약 450bp의 PCR 증폭산물이 확인되었다(자료 미제시). PCR 증폭산물의 DNA 염기 서열 분석 결과를 토대로 분자생물학적 계통수를 작성하여 정확한 종을 동정하였으며, 그 결과는 Fig. 1과 같다.
Fig. 1
Bayesian tree inferred from cytochrome c oxidase subunit I (COI) gene sequences dataset of mitochonfrial DNA with GTR+G substitution model between ‘Schachtii’ group species of Heterodera. Posterior probability more than 50% is given on appropriate clades. The newly obtained sequences from Jeongseon, Korea are indicated in bold.

BC115, YS224, DA142 시료에서 검출된 씨스트선충의 종내변 이율은 0%로 나타났다. NCBI에 기등록된 근연종 H. glycines (KC172914)와 종간변이율은 0.3%, H. schachtii (KC172918)와는 7.2%, H. ciceri (KC172919)와는 10.1%, H. trifolii (KT163239) 와는 9.9%로 나타났다. 이에 따라 BC115, YS224, DA142 시료에서 검출된 씨스트선충은 콩씨스트선충(H. glycines)으로 동정되었다. 한편, PM001, GC147, GC408 시료에서 검출된 씨스트선충의 종내변이율(Intraspecific)은 0%로 나타났다. NCBI 에 등록되어 있는 근연종과의 종간변이율(Interspecific) 분석 결과, H. schachtii (KC172918)와 종간변이율은 0%, H. glycines (KC172914)와는 7.7%, H. ciceri (KC172919)와는 13.2%, H. trifolii (KC163239)와는 12.3%로 나타났다. 분석 결과에 따라 PM001, GC147, GC408 시료에서 검출된 씨스트선충은 사탕무씨스트선충(H. schachtii)으로 동정되었다. Vovlas 등(2015)의 연구에서 mtDNA COI 유전자의 분자생물학적 계통수 분석 결과, 사탕무씨스트선충(H. schachtii)과 콩씨스트선충(H. glycines) 등 Heterodera 속 ‘Schachtii’ 그룹의 씨스트선충 5종을 높은 종 동정 해상력(Resolving power)으로 구별하였다. 본 연구결과에서도 mtDNA COI 유전자 영역을 통해 2종의 Heterodera 속 씨스트선충을 명확히 구별할 수 있었다.
PCR-RFLP를 이용한 씨스트선충 2종의 판별
분자생물학적 계통수 분석을 통해 동정된 사탕무씨스트선충(H. schachtii) GC147 시료와 콩씨스트선충(H. glycines) YS224 시료의 PCR 증폭산물을 표준집단으로 선발하여 8종류의 제한효소 처리에 따른 두 종간의 절편길이다형성(RFLP)을 비교하고자 하였다 (Fig. 2, Table 2). 그 결과, RsaI 제한효소 처리 시 사탕무씨스트선충(H. schachtii)은 169, 279 bp 2개의 DNA 밴드양상을 나타냈으며, 콩씨스트선충(H. glycines)은 448 bp의 단일밴드만 확 인되었다. HinfI 처리시 콩씨스트선충은 50, 398 bp의 DNA 밴드 양상을 나타냈으나, 사탕무씨스트선충은 448 bp의 단일밴드만 확인되었다.
Fig. 2
Restriction fragment length polymorphism (RFLP) of Heterodera schachtii and H. glycines detected from chinese cabbage fields in Jeongseon, Korea. (A) H. schachtii, GC147, (B) H. glycines, YS224, Lane M: 100 bp DNA ladder, 1: PstI, 2: VspI, 3: AlwI, 4: RsaI, 5: MvaI, 6: EcoRI, 7: Eco72I, 8: HinfI.
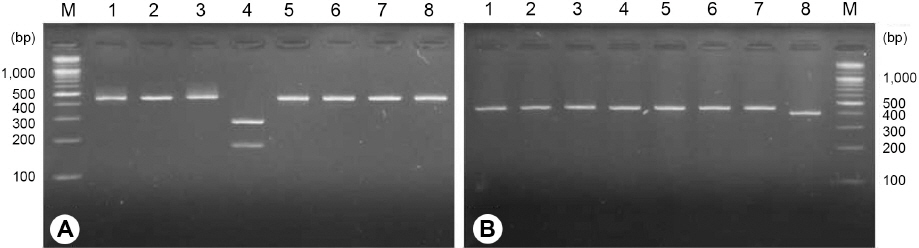
Table 2
Restriction fragment length polymorphism of 2 cyst forming nematodes based on difference of mtDNA COI gene sequences
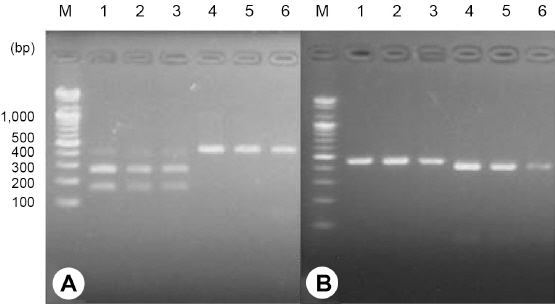
사탕무씨스트선충과 콩씨스트선충의 RFLP 분석 결과를 토대로, 다른 포장에서 검출된 사탕무씨스트선충과 콩씨스트 선충에 RsaI과 HinfI 제한효소 처리시에도 동일한 결과가 나타나는지 여부를 확인하였다(Fig. 3). 사탕무씨스트선충(H. schachtii)으로 동정되었던 PM001, GC408 포장에서 검출된 씨스트선충의 PCR 증폭산물에 RsaI 처리시 GC147번 시료(H. schachtii)와 동일하게 169, 279 bp 2개의 DNA 밴드양상을 나타냈으며, HinfI 처리시에는 448 bp의 단일밴드를 형성하였다. 콩씨스트선충(H. glycines)으로 동정되었던 BC115, DA142 포장에서 검출된 씨스트선충의 PCR 증폭산물에 RsaI 처리시 YS224번 시료(H. glycines)와 동일하게 448 bp의 단일밴드를 형성하였으며, HinfI 처리시에는 50, 398 bp 2개의 DNA 밴드양상을 나타냈다. 따라서, 사탕무씨스트선충과 콩씨스트선충 2종의 mtDNA COI 영역 PCR 산물에 RsaI과 HinfI을 처리하면 2종을 구별할 수 있을 것으로 생각된다.
Fig. 3
Restriction fragment length polymorphism of cyst forming nematodes detected from 6 chinese cabbage fields in Samcheok and Jeongseon. (A) RsaI treatment (B) HinfI treatment, lane M: 100 bp DNA ladder, 1-3: Heterodera schachtii, 1: GC147, 2: GC408, 3: PM001, 4-6: H. glycines, 4: YS224, 5: DA142, 6: BC115.
Choi (2000)와 The Korean Society of Plant Pathology (2009) 에 따르면, 국내 보고된 Heterodera 속 씨스트선충은 벼씨스트선충(H. elachista), 왕벼씨스트선충(H. oryzae), 콩씨스트선충 (H. glycines), 사탕무씨스트선충(H. schachtii) 4종이 있는 것으로 알려져 있다. 벼씨스트선충의 경우 NCBI에 기등록되어 있는 KC618473의 DNA 염기서열 정보를 이용하여 Webcutter 2.0 (http://rna.lundberg.gu.se/cgi-bin/cutter2)에서 제한효소 인식부위를 확인한 결과, RsaI과 HinfI 제한효소의 인식부위가 없는 것으로 나타났다. 이에 따라, 벼씨스트선충도 RsaI과 HinfI 제한효소 처리를 통해 사탕무씨스트선충 및 콩씨스트선충과 종 구분이 가능할 것으로 판단된다. 왕벼씨스트선충은 mtDNA COI 유전자의 염기서열 정보가 없어 RFLP 결과 비교에 어려움이 있었다. 향후 연구에서는 왕벼씨스트선충과 벼씨스트선충을 포함한 국내 분포하는 Heterodera 속 씨스트선충 4종에 대한 PCR-RFLP 추가 연구가 수행되어야 할 것이다.
종 특이 프라이머를 이용한 씨스트선충 2종의 판별
고랭지 배추 재배지에서 검출된 사탕무씨스트선충(H. schachtii)과 콩씨스트선충(H. glycines)의 종 판별을 위해 본 연구에서 설계한 특이 프라이머 세트(#JBS1, #JBG1, #JB3R)를 이용하여 PCR을 수행하였다(Fig. 4). 콩씨스트선충은 339 bp의 단일 증폭산물이 확인되었으며, 삼척 1개(BC115)와 정선 2개(YS224, DA142)의 포장에서 검출된 콩씨스트선충에서도 모두 동일한 결과를 얻었다. 반면, 사탕무씨스트선충은 예상했던 277 bp 증폭산물 이외에도 339 bp의 증폭산물이 확인되었으며, 삼척 1개(PM001)와 정선 2개(GC147, GC408)의 포장에서 검출된 사탕무씨스트선충에서도 모두 동일한 결과를 나타냈다. 본 연구결과에서 사 탕무씨스트선충의 특이 프라이머 #JBS1은 콩씨스트선충 DNA 주형에 분명히 프라이머로 작동하지 않았다. 그러나, 콩씨스트선충 특이프라이머는 #JBG1의 3’ 말단 염기와 사탕무씨스트선충 DNA 주형의 #JBG1 프라이머 인식서열 3’ 말단 염기가 시토신(cytosine)과 티민(thymine)으로 서로 불일치했으나, #JBG1 프라이머가 작동하여 사탕무씨스트선충 DNA도 증폭되었다. Kwok 등(1990)과 Ayyadevara 등(2000)에 의하면 프라이머와 DNA 주형의 3’ 말단 염기가 서로 불일치하면 PCR 효율이 최대 1/100배 정도로 크게 감소하는 것으로 알려져 있다. 그러나, 본 연구결과에서는 콩씨스트선충의 특이프라이머와 사탕무씨스트선충 DNA 주형의 3’ 말단 염기가 서로 불일치하였지만 PCR 효율에는 크게 영향을 미치지 않아 사탕무씨스트선충의 DNA 단편도 증폭된 것으로 생각된다. 한편, 고구마뿌리혹선충, 땅콩 뿌리혹선충, 당근뿌리혹선충, 딸기뿌리썩이선충과 사과뿌리썩이선충에서는 PCR 증폭산물이 검출되지 않았으며, 사탕무씨스트선충과 콩씨스트선충의 DNA 밴드 양상을 통해 두 종을 구분할 수 있을 것으로 생각된다.
Fig. 4
PCR result with species specific primer set for Heterodera schachtii and H. glycines. M: 100 bp DNA ladder, 1-3: H. schachtii, 1: GC147, 2: GC408, 3: PM001, 4-6: H. glycines, 4: YS224, 5: DA142, 6: BC115, 7: M. incognita, 8: M. arenaria, 9: M. hapla, 10: P. penetrans, 11: P. vulnus, 12: negative control.

GC147 시료의 사탕무씨스트선충과 YS224 시료의 콩씨스트선충 DNA 농도에 따른 종 특이 프라이머 세트의 PCR 민감도 검정 결과, GC147 시료의 사탕무씨스트선충은 1반응물(reaction) 당 100 ng, 10 ng, 1 ng과 100 pg의 농도에서 PCR 증폭산물이 검출되었다(Fig. 5). 반면, YS224 시료의 콩씨스트선충은 1반응 물 당 100 ng, 10 ng과 1 ng의 농도에서만 PCR 증폭산물이 검출되었다(Fig. 5). 본 과정을 3회 반복(biological replicates)하여도 동일한 결과를 얻었으며(자료 미제시), 사탕무씨스트선충과 콩씨스트선충 종 판별을 위한 PCR의 DNA 적정 농도는 1반응물 당 10 ng 정도로 생각된다. Subbotin 등(2001)과 Amiri 등 (2002)의 연구에서도 사탕무씨스트선충과 콩씨스트선충 ITS rRNA 영역에서의 염기서열 차이를 이용하여 종 특이 프라이머 개발에 관한 연구를 수행한 바 있으나, 민감도에 대한 연구는 수행되지 않아 특이 프라이머 간의 민감도 비교에는 어려움이 있었다. 한편, 국내에는 이들 2종 이외에도 벼씨스트선충 (H. elachista)과 왕벼씨스트선충(H. oryzae), 대나무에서 발견 된 씨스트선충(H. koreana) 등이 분포하고 있는 것으로 알려져 있다. 향후 연구에서는 PCR-RFLP와 마찬가지로 국내 분포하는 Heterodera 속 씨스트선충의 다양한 종을 확보하고, 신속하고 정확한 종 동정을 위해 Multiplex PCR 기법과 같은 진단 기술의 추가적인 연구가 필요할 것으로 생각된다.
Fig. 5
PCR sensitivity of specific primer set for Heterodera schachtii and H. glycines DNAs with different concentration. M : 100 bp DNA ladder, 1-6: H. schachtii (GC147) 1: 100 ng, 2: 10 ng, 3: 1 ng, 4: 100 pg, 5: 10 pg, 6: 1 pg, 7-12: H. glycines (YS224) 7: 100 ng, 8: 10 ng, 9: 1 ng, 10: 100 pg, 11: 10 pg, 12: 1.

요약
강원도 고랭지배추 포장에서 검출된 사탕무씨스트선충(H. schachtii)과 콩씨스트선충(H. glycines)을 구별할 수 있는 신속 진단법을 개발하고자 하였다. 이를 위해 mtDNA COI 유전자 영역의 계통분석으로 동정된 사탕무씨스트선충 GC147, GC408, PM001 개체군과 콩씨스트선충 YS224, DA142, BC115 개체군을 대상으로 PCR-RFLP와 본 연구에서 개발한 특이 프라이머 세트를 이용한 PCR을 수행하였다. 사탕무씨스트선충과 콩씨스트선충 각각 3개 개체군의 mtDNA COI 영역 PCR 증폭산물에 8종류의 제한효소를 처리하여 DNA 절편길이다형성을 확인하였으며, 2종류의 제한효소 RsaI과 HinfI을 처리하면 DNA 밴드 양상의 차이로 사탕무씨스트선충과 콩씨스트선충 두 종을 구별할 수 있었다. 또한, 본 연구에서 개발한 프라이머 세트(JBS1, JBG1과 JB3R)는 사탕무씨스트선충 mtDNA COI 영역의 277과 339 bp, 콩씨스트선충의 339 bp의 특정 DNA 단편을 증폭시켰으며, 뿌리혹선충 3종과 뿌리썩이선충 2종의 식물기생선충은 증폭시키지 않았다. 따라서, 본 연구에서 개발한 프라이머 세트를 이용하여 PCR을 수행하면 사탕무씨스트선충과 콩씨스트선충을 구별할 수 있었다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






