

서론
딸기의 총 생산액은 약 1조 3천억 원으로 미곡 다음으로 큰 시장을 형성하고 있으며, 농가에서도 안정적이고 고소득을 낼 수 있는 작물로 여기고 있다(MAFRA, 2015). 대부분의 딸기 시설재배 농가들이 연작을 하고 있으며, 이에 따라 식물기생선충에 의한 피해가 지속적으로 나타나고 있다(Kim 등, 2013).
딸기에 피해를 주는 식물기생선충으로는 뿌리혹선충(Meloidogyne), 뿌리썩이선충(Pratylenchus), 잎선충(Aphelenchoides), 창선충(Xiphinema), 바늘선충(Longidorus), 주름선충(Criconematoides) 등이 알려져 있다(Brown 등, 1993; Esnard와 Zukerman, 1998). 또한, 국내 딸기 노지재배지와 시설재배지를 대상으로 토양 내 식물기생선충 감염현황을 조사한 결과에서도 뿌리혹선충과 뿌리썩이선충이 주요 문제 선충으로 나타난 바 있다(Choi, 1977; Ko 등, 2016). 이러한 식물기생선충의 종 동정은 형태적 특징과 각 형태 부위별 길이, 폭 등의 평균 측정치가 주로 이용되어 왔으나(Powers, 2004), 최근에는 28S rRNA D2뺻3 부위 유전자나 mitochondrial DNA cytochrome c oxidase subunit II (COII) 부위 유전자와 같은 특정 유전자 부위를 대상으로 한 중합효소연쇄반응(PCR), 제한효소절편길이다형성(restriction fragment length polymorphism) 등과 같은 분자생물학적 기법이 널리 이용되고 있다.
식물기생선충은 이들이 감염된 토양이 묻은 작업화, 농기계 및 관개수 등을 통해 인근 포장으로 확산될 수 있는 것으로 알려져 있다(Castillo 등, 2007). 특히, 딸기는 모주(mother plant)의 런너(runner)에 의해 생산되는 자묘(daughter plant)를 모종으로 이용한다. 딸기 자묘가 식물기생선충이 감염된 토양과 접촉하면 식물기생선충은 딸기의 본포장으로 확산될 수 있으며, 이를 막기 위해서는 식물기생선충이 감염되지 않은 토양에서 건전한 육묘를 생산하는 것이 중요하다. 하지만 국내에서는 아직까지 딸기 육묘장에서 문제될 수 있는 식물기생선충의 감염현황, 피해와 확산 등에 대한 연구가 수행되지 않았다.
이에 따라, 본 연구에서는 딸기 육묘장의 지역, 육묘형태 및 육묘에 이용되는 토양종류별 식물기생선충의 감염현황을 조사하고, 문제 선충의 종을 동정하며, 이들이 자묘를 통해 인근지역이나 타지역의 딸기 본포장으로 확산될 수 있는지 여부를 조사하고자 하였다.
재료 및 방법
딸기 육묘장 토양 및 자묘 채취와 선충분리
2016년 6월 24일부터 9월 1일까지 충남의 2개 지역(논산, 부여), 전남의 3개 지역(담양, 곡성, 순천)과 경남의 2개 지역(진주, 밀양) 총 7개 지역의 딸기 육묘장으로부터 117점의 토양시료를 채취하였다. 토양시료는 육묘장의 지역, 육묘형태 및 육묘토양 종류를 기준으로 구분하여 채취하였다. 모든 시료는 분석에 사용하기 전까지 10°C 저온창고에 보관하였다.
채취한 토양으로부터 선충을 분리하기 위해 토양을 골고루 섞어 플라스틱 비커를 이용하여 300 cm3를 정량하고, 수돗물이 들어있는 플라스틱 통에 넣어 현탁액을 만들었다. 토양현탁액을 20 mesh와 400 mesh 체(sieve)를 이용하여 거른 다음 변형시킨 Baermann 깔때기에 올려놓고 실험실 조건에서 48시간 동안 배양하였다(Barker 등, 1985; Kim 등, 2014). 48시간 경과 후, 변형시킨 Baermann 깔때기의 고무튜브에 모인 선충을 15 ml 플라스틱 튜브를 이용하여 회수하였다. 분리된 선충은 실체현미경(MZ12; Leica, Wetzlar, Germany)을 이용하여 50배율로 관찰하면서 식물기생선충의 종류와 밀도를 조사하였다.
딸기 육묘장 주요 문제 선충의 DNA 추출과 PCR
국내 딸기 육묘장의 주요 문제 선충 종 동정을 위해 부여 1개, 담양 1개와 순천 3개 포장으로부터 분리된 뿌리썩이선충과 담양 1개, 순천 1개 포장으로부터 분리된 뿌리혹선충을 각각 3×10마리씩 표준집단으로 선발하여 Iwahori 등(2000)의 방법을 통해 genomic DNA를 추출하였다(Table 1). 슬라이드글라스 위에 멸균수를 한 방울 떨어뜨리고 뿌리썩이선충 또는 뿌리혹선충을 건져 넣은 다음, 작게 자른 필터페이퍼 조각으로 선충을 짓눌렀다. 이때, 체벽이 파쇄되면서 나오는 세포성분을 필터페이퍼에 묻혀 15 µl의 Proteinase K solution (2 M KCl2, 10 mM Tris-HCl, 1 M MgCl2, 10% [w/v] Triton-X 100, 20.6 mg/ml Proteinase K)이 들어있는 PCR tube에 넣고 60°C에서 30분, 94°C에서 10분간 배양시켜 DNA를 추출하였다.
Table 1
The origin of nematodes used in phylogenetic analysis
뿌리썩이선충의 PCR은 28S rRNA D2뺻3 expansion segments 부위 유전자에 대한 universal primer인 #D2A (forward primer, 5’-ACAAGTACCGTGAGGGAAAGTTG-3’)와 #D3B (reverse primer, 5’-TCGGAAGGAACCAGCTACTA-3’)를 이용하여 수행하였다(Nunn, 1992). 뿌리혹선충의 PCR은 COII 부위 유전자에 대한 universal primer인 #C2F3 (forward primer, 5’-GGTCAATGTTCAGAAATTTGTGG-3’)와 #1108 (reverse primer, 5’-TACCTTTGACCAATCACGCT-3’)를 이용하여 수행하였다(Powers와 Harris, 1993). PCR을 위해 template DNA 4 µl, PCR premix (HelixAmp™ Ready-2X-Go Series; Nanohelix, Daejeon, Korea) 15 µl, forward와 reverse primer 각각 0.5 µl와 triple distilled water 30 µl를 0.2 ml centrifuge 튜브에 넣고 혼합하였다. 혼합한 0.2 ml의 튜브를 PCR cycler (PTC-200; MJ Research, Alameda, CA, USA)를 이용하여 뿌리썩이선충은 94°C에서 1분간 denaturation, 55°C에서 40초간 annealing, 72°C에서 1분간 extension 과정을 35회 반복하였으며, 뿌리혹선충은 94°C에서 1분간 denaturation, 48°C에서 1분간 annealing, 68°C에서 3분간 extension 과정을 35회 반복하였다. PCR 결과는 전기영동장치(Mupid eXu; ADVANCE, Tokyo, Japan)를 이용하여 1% 아가로오스겔, 1× TAE buffer (0.04 M Tris-acetate, 0.001 M EDTA)에서 100 V, 25 mA로 25분간 전기영동한 다음, UV장치(UVCI-1100; Major Science, New Taipei City, Taiwan)에서 확인하였다.
분자생물학적 계통수 및 종내종간· 변이율 분석
뿌리썩이선충과 뿌리혹선충의 PCR 증폭산물은 PCR Purification Kit (Macherey-Nagel, Düen, Germany)를 이용하여 정제하였으며, DNA 염기서열은 (주)제노텍 유전체분석센터에서 Sanger dideoxy sequencing 기반 장비(ABI PRISM 3730xl DNA sequencer; Applied Biosystems, Foster City, CA, USA)를 이용하여 분석하였다. 분석 결과로부터 Chromas Lite 2.0, EditSeq 5.05와 SeqMan 5.05 프로그램을 이용하여 DNA 염기서열을 결정하였으며, 미국국립생물정보센터(National Center for Biotechnology Information, NCBI)에 기등록된 데이터들과 함께 Clustal X 1.83의 기본값으로 정렬하였다. 분자생물학적 계통수는 Mrbayes 3.2.6의 General Time Reversible substitution model (GTR+G)을 이용하여 작성하였고(Vovlas 등, 2015), Dendroscope 3.5.7을 이용하여 편집하였다. 뿌리썩이선충의 외집단분류군(outgroup)은 Palomares-Rius 등(2010)의 연구결과를 참고하였으며, 뿌리혹선충의 외집단분류군은 Humphreys-Pereira 등(2014)의 연구결과를 참고하였다. 종내 · 종간 변이율 분석에는 MEGA 6.06의 distance 분석 도구를 이용하였다.
딸기 자묘를 통한 뿌리썩이선충의 포장 간 분산 여부 검정
2016년 8월 9일, 순천의 사과뿌리썩이선충(Pratylenchus vulnus)이 감염된 딸기 노지 토경육묘장으로부터 생산된 자묘를 3포기 채취하였다. 채취한 자묘 뿌리를 수돗물로 깨끗이 씻은 다음 1,000 cm3의 멸균토양이 들어있는 토분(직경 13 cm)에 2016년 8월 11일 정식하였다. 정식 33일 후, 토양 내 뿌리썩이선충 밀도를 확인하기 위해 20 mesh와 400 mesh 체를 이용한 체법과 Baermann 깔때기법으로 선충을 분리하였다. 실체현미경(MZ12)을 이용하여 분리한 선충 시료에서 뿌리썩이선충의 존재 여부와 밀도를 확인하였다.
결과 및 고찰
딸기 육묘장의 식물기생선충 감염현황
2016년 6월부터 9월까지 국내 딸기 육묘장 117개 포장의 토양시료의 식물기생선충 감염현황을 조사한 결과, 딸기 육묘장의 토양에서는 뿌리혹선충(Meloidogyne spp.), 뿌리썩이선충(Pratylenchus spp.)과 참선충(Tylenchidae)이 검출되었다(Table 2). 주요 검출 선충 가운데 참선충은 딸기에 피해를 일으키지 않는 선충으로 알려져 있기 때문에(Choi, 1977) 딸기 육묘장에서 가장 문제되는 선충은 뿌리썩이선충과 뿌리혹선충으로 생각된다. 이러한 결과는 시설재배 주산단지 포장에서 뿌리썩이선충과 뿌리혹선충이 주요 문제 선충으로 나타난 연구 결과와 일치하였다(Ko 등, 2016). 뿌리썩이선충의 지역별 검출빈도는 순천과 논산이 33%와 20%로 가장 높았으며, 뿌리혹선충은 담양과 순천에서 10%와 13%의 비교적 낮은 빈도로 검출되었다. 그러나 딸기 주요 기생선충이 자묘를 통해 본포장으로 확산될 수 있음을 고려하였을 때, 결코 낮지 않은 검출빈도로 생각된다. 한편, 딸기의 문제 선충으로 알려져 있는 잎선충(Aphelenchoides spp.)의 발생 조사를 위해 117개 포장 가운데 피해가 의심되는 29개 포장에서 자묘를 채취하여 잎선충 감염 여부를 조사하였으나 검출되지 않았다(data not shown). 이는 잎선충이 감염되지 않은 조직배양묘의 보급이나 피해 의심주 발생 시 농가에서 조기 제거하기 때문에 검출되지 않았을 것으로 생각된다.
Table 2
The incidence of plant-parasitic nematodes in strawberry nursery
| Location | No. of soil samples | % of nematode infestation (nematode density*) | ||
|---|---|---|---|---|
|
|
||||
| Meloidogyne | Pratylenchus | Tylenchidae | ||
| Nonsan | 25 | 0 | 20 (8.4±7.44) | 36 (6.5±5.94) |
| Buyeo | 12 | 0 | 8 (6†) | 8 (77) |
| Damyang | 21 | 10 (7.5±7.78) | 10 (24.5±26.16) | 5 (4) |
| Gokseong | 12 | 0 | 0 | 17 (4±2.83) |
| Suncheon | 15 | 13 (4.5±3.54) | 33 (63.2±89.15) | 20 (15±23.39) |
| Jinju | 22 | 0 | 0 | 0 |
| Miryang | 10 | 0 | 0 | 20 (23.5±16.26) |
| Total | 117 | 3 (6.0±5.23) | 11 (31.8±58.55) | 15 (14.4±19.33) |
딸기의 육묘형태와 토양종류별 식물기생선충 감염현황
국내 딸기 육묘장 117개 포장을 대상으로 육묘 형태와 토양의 종류별 식물기생선충의 발생현황을 조사한 결과는 Table 3과 같다. 노지(field) 또는 비닐하우스(plastic house)에서 밭토양(upland soil)을 이용하여 토경육묘를 했을 때, 딸기의 주요 기생선충으로 알려진 뿌리썩이선충과 뿌리혹선충(Ko 등, 2016)의 검출빈도는 노지에서 17%와 2%, 비닐하우스에서 21%와 13%로 다른 육묘방법에 비해 상대적으로 높게 나타났다. 비닐하우스에서 화강토(granite soil)를 이용하여 포트육묘를 했을 때는 뿌리썩이선충이 5%의 낮은 빈도로 검출되었다. 이와 달리, 노지에서 화강토를 이용하거나 비닐하우스에서 상토(bed soil)를 이용하여 포트육묘를 했을 때 뿌리썩이선충과 뿌리혹선충 등 어떠한 식물기생선충도 검출되지 않았다. 하지만 노지와 비닐하우스의 포트육묘에 이용되는 화강토는 동일하므로 화강토를 이용한 노지 포트육묘에서도 식물기생선충이 검출될 수 있는 가능성을 배제할 수 없다. 한편, 국내에서는 딸기 육묘에 있어 잎선충의 피해를 예방하기 위해 모주 정식전 살선충제 침지처리를 통한 방제 효과에 대한 연구 이외에는 모주재배지의 선충 관리방안에 대한 연구가 수행되지 않았다(Kim 등, 2008). 이에 따라, 비닐하우스에서 상토를 이용하여 포트육묘를 하면 잎선충을 제외한 토양 내 서식하는 주요 기생선충의 감염을 사전에 예방할 수 있는 친환경적인 선충 관리방안으로 생각된다.
Table 3
The incidence of plant-parasitic nematodes in strawberry nurseries with different nursery and soil types
| Nursery type | Soil type | No. of samples | % of nematode infestation (nematode density*) | |||
|---|---|---|---|---|---|---|
|
|
||||||
| Meloidogyne | Pratylenchus | Aphelenchoides | Tylenchidae | |||
| Field | Upland soil | 41 | 2 (7.3±5.51) | 17 (33.9±60.62) | 0 | 39 (15.0±19.75) |
| Plastic house | Upland soil | 24 | 13 (6.0±5.23) | 21 (31.8±58.55) | 0 | 8 (14.4±19.33) |
| Pot on field | Granite soil | 11 | 0 | 0 | 0 | 0 |
| Pot in plastic house | Granite soil | 20 | 0 | 5 (2†) | 0 | 0 |
| Pot in plastic house | Bed soil | 21 | 0 | 0 | 0 | 0 |
딸기 육묘장 문제 선충의 종 동정
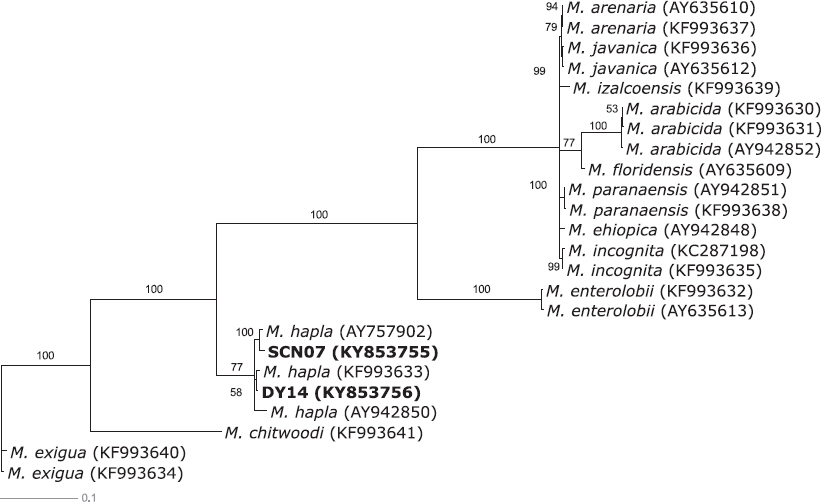
딸기 육묘장의 문제 선충으로 나타난 뿌리썩이선충과 뿌리혹선충의 종을 확인하기 위해 유전자 분석을 수행하였다(Fig. 1). 유전자 분석을 위해 뿌리썩이선충은 28S rRNA 유전자 부위 마커를, 뿌리혹선충은 COII 유전자 부위 마커를 선발하여 PCR을 수행하였다. 그 결과 뿌리썩이선충의 모든 시료로부터 약 750 bp의 증폭산물을 얻었으며, 뿌리혹선충의 모든 시료로부터 약 500 bp의 증폭산물을 얻었다(data not shown). DNA 염기서열을 기반으로 분자생물학적 계통분석을 위해 각 선충 표준집단의 PCR 증폭산물에 대한 DNA 염기서열 분석을 수행하였으며, 이를 통해 작성한 분자생물학적 계통수는 Fig. 1, 2와 같다. 부여 1개, 담양 1개와 순천 3개 포장에서 각각 검출된 뿌리썩이선충과 담양 1개, 순천 1개 포장에서 각각 검출된 뿌리혹선충의 분자생물학적 계통분석 결과, 딸기뿌리썩이선충(Pratylenchus penetrans)과 사과뿌리썩이선충(P. vulnus) 2종의 뿌리썩이선충과 당근뿌리혹선충(Meloidogyne hapla) 1종의 뿌리혹선충이 확인되었다.
Fig. 1
The bayesian tree inferred from 28S rRNA expansion segments D2-D3 region sequences with general time reversible substitution model (GTR+G) of Pratylenchus. Newly obtained sequences of Pratylenchus species indicated in bold.
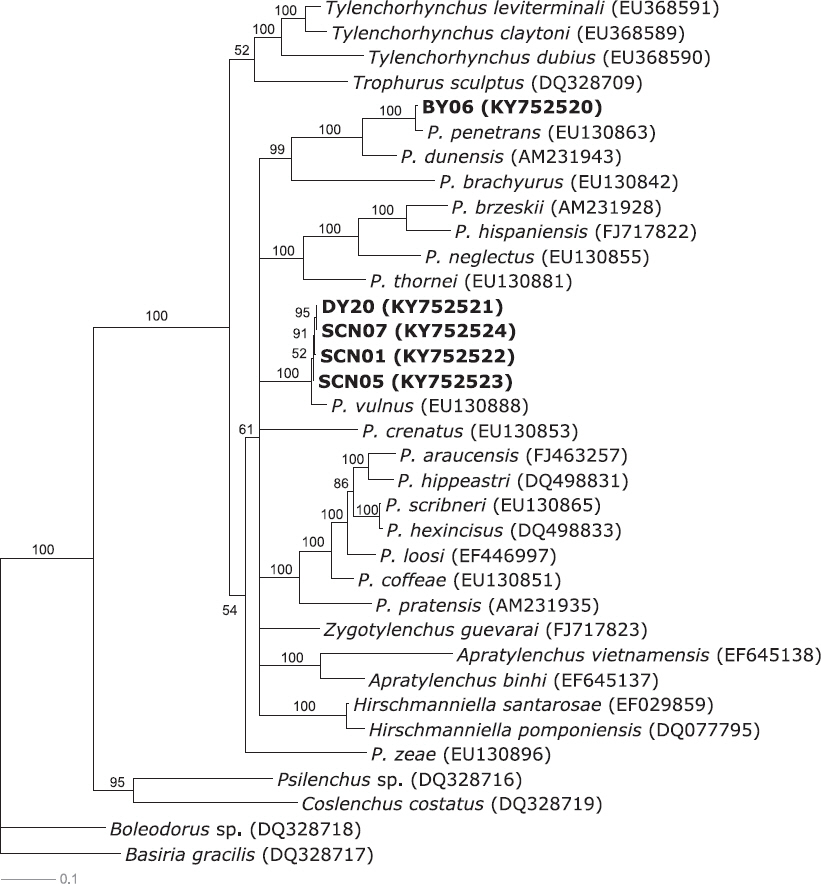
Fig. 2
The bayesian tree inferred from cytochrome c oxidase subunit II (COII) region sequences with general time reversible substitution model (GTR+G) of Meloidogyne. Newly obtained sequences of Meloidogyne species indicated in bold.
정확한 종 동정을 위해 MEGA 6.06 프로그램을 이용하여 distance 분석을 수행하였다. 딸기뿌리썩이선충 clade로 분류된 BY06 개체군(population)을 NCBI에 기등록된 P. penetrans (EU130863)와 종내 변이율을 분석한 결과, P. penetrans (EU130863)와는 1.9%의 종내 변이율을 나타냈다. 근연종과의 종간 변이율 분석에는 P. penetrans (EU130863)와 Pratylenchus dunensis (AM231943)를 이용하였으며, P. penetrans (EU130863)와는 1.9%, P. dunensis (AM231943)와는 12.6%의 종간 변이율을 나타냈다. 이러한 결과에 따라, 부여 1개 포장에서 검출된 뿌리썩이선충은 딸기뿌리썩이선충(P. penetrans)으로 동정되었다.
사과뿌리썩이선충 clade로 분류된 DY20, SCN01, SCN05와 SCN07 국내 개체군들 간의 종내 변이율을 분석한 결과 0.2%로 나타났다. 근연종과의 종간 변이율 분석에는 P. vulnus (EU130888), Pratylenchus crenatus (EU130853)와 Pratylenchus pratensis (AM231935)를 이용하였으며, P. vulnus (EU130888)와는 2.8%, P. crenatus (EU130853)와는 20.3%, P. pratensis (AM231935)와는 22.6%의 종간 변이율을 나타냈다. 이에 따라, 담양 1개와 순천 3개 포장에서 검출된 뿌리썩이선충은 사과뿌리썩이선충으로 동정되었다.
당근뿌리혹선충 clade로 분류된 SCN07과 DY14 국내 개체군들 간의 종내 변이율을 분석한 결과 1.4%로 나타났다. 근연종과의 종간 변이율 분석에는 M. hapla (AY757902, KF993633), Meloidogyne chitwoodi (KF993641)와 M. enterolobii (KF993632)를 이용하였다. SCN07, DY14와 M. hapla (AY757902)와는 2.5%, M. hapla (KF993633)와는 1.0%, M. chitwoodi (KF993641)와는 32.9%, M. enterolobii (KF993632)와는 23.1%의 종간 변이율을 나타냈다. 이를 통해 담양 1개와 순천 1개 포장에서 검출된 뿌리혹선충은 당근뿌리혹선충(M. hapla)으로 동정되었다.
사과뿌리썩이선충과 딸기뿌리썩이선충은 국내 딸기재배지 일부 지역의 선충 조사 결과에서 검출이 확인된 바 있으며(Choi, 1977; Park 등, 2005), 사과뿌리썩이선충과 당근뿌리혹선충은 진주지역 딸기 시설재배 포장의 문제 선충으로도 잘 알려져 있다(Ko 등, 2016). 본 연구 결과에 따라, 딸기 육묘장에서 자묘를 생산함에 있어 딸기뿌리썩이선충, 사과뿌리썩이선충과 당근뿌리혹선충의 피해가 문제될 것으로 생각되며, 육묘 전 이들의 감염 여부 확인 및 방제 대책을 마련하는 것이 필요할 것으로 생각된다.
딸기 자묘를 통한 식물기생선충의 포장 간 분산
순천에서 채취한 사과뿌리썩이선충이 감염된 딸기 자묘를 멸균토양에 정식했을 때 33% 빈도로 사과뿌리썩이선충의 감염이 확인되었다. Castillo 등(2007)에 의하면 뿌리썩이선충은 스스로 이동할 수 있는 거리가 제한되어 있기 때문에 뿌리썩이선충 스스로 다른 포장으로의 확산은 빠르게 일어날 수 없다고 하였다. 하지만 뿌리썩이선충이 감염된 토양이 묻은 농기구나 신발을 통해서 빠르게 확산될 수 있고, 관개수를 통해서 먼 거리를 이동할 수 있다고 하였다. 사탕무씨스트선충(Heterodera schachtii)도 작업화나 작업차량에 묻은 토양을 통해 인근 포장으로 확산될 수 있으며, 강우 시 유거수를 통해서도 확산될 수 있다고 알려진 바 있다(Kwon 등, 2016). 딸기의 경우 런너를 통해 생성되는 자묘를 모종으로 이용하기 때문에 딸기 주요 기생선충이 자묘 뿌리에 묻은 토양을 통해서 확산될 가능성이 있을 것으로 생각된다. 그러나 본 연구에서는 뿌리썩이선충이 감염된 토양을 제거한 자묘를 통해서도 이들 선충이 다른 포장으로 확산될 수 있는지 여부를 확인하고자 하였다. 그 결과, 토양이 묻지 않은 자묘를 통해서도 뿌리썩이선충이 확산될 수 있음을 확인하였다. 뿌리혹선충도 마찬가지로 딸기 뿌리에 혹과 난낭을 형성하므로 자묘를 통해서 다른 포장으로 확산될 수 있을 것으로 생각된다. 이에 따라 딸기 육묘 시 모주를 정식하기 전 관할지역 시군농업기술센터나 도농업기술원을 통해 딸기 육묘예정지에 뿌리썩이선충이나 뿌리혹선충과 같은 주요 문제 선충이 감염되었는지를 확인하고 육묘에 들어가는 것이 좋을 것으로 생각된다.
요약
2016년 6월부터 9월까지 국내 딸기 육묘장 117개 포장의 토양으로부터 식물기생선충의 토양 내 감염현황을 조사하였다. 그 결과 뿌리썩이선충(Pratylenchus)과 뿌리혹선충(Meloidogyne)이 11%와 3%로 검출되어 딸기 육묘장의 문제 선충으로 나타났다. 육묘 형태별로는 토경육묘에서 뿌리썩이선충과 뿌리혹선충의 발생빈도가 다른 육묘방법에 비해 높았고, 상토를 이용한 비가림 포트육묘에서는 식물기생선충이 검출되지 않았다. 뿌리썩이선충은 딸기뿌리썩이선충(P. penetrans)과 사과뿌리썩이선충(P. vulnus) 2종이 확인되었으며, 뿌리혹선충은 당근뿌리혹선충(M. hapla) 1종이 확인되었다. 사과뿌리썩이선충은 딸기 자묘를 통해 식물기생선충이 감염되지 않은 건전한 토양으로의 확산이 확인되었다(33%).
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






