

서론
전 세계적으로 생산되는 주요 농작물이 식물 병원성 해충 및 선충에 의해 그 수확량이 12%-40%까지 감소하고 콜로라도감자잎벌레(Leptinotarsa decemlineata), 옥수수뿌리벌레(Diabrotica virgifera), 배추좀나방(Plutella xylostella), 은잎가루이(Bemisia argentifolii), 알락수염노린재(Dolycoris baccarum) 등의 해충에 의한 피해액은 연간 300억 달러에 달하는 것으로 보고되었다(FAO, WFP and IFAD, 2012). 식물기생성 선충은 대표적으로 뿌리혹선충, 포낭선충, 뿌리썩이선충 등이 보고되어 있으며(Degenkolb와 Vilcinskas, 2016; Jansson과 López-Llorca, 2004), 전 세계적으로 이로 인한 농작물의 수확량이 해마다 평균 12% 감소하고 피해액이 1천 300억 달러에 이른다(Becker, 2014; Degenkolb와 Vilcinskas, 2016). 이러한 농작물의 손실을 줄이기 위해 세계적으로 연간 약 30억 킬로그램 가량의 살충제를 사용하는 것으로 알려졌다(Pimentel, 2009). 이러한 식물병원성 해충과 선충을 방제하기 위하여 사용되는 화학물질로는 유기인산 화합물(살충제), 포스티아제이트(살선충제), 카바메이트계 화합물(살충제), 다조메트(살선충제) 등의 화학 약제를 사용하고 있다. 하지만 화학적 방제제는 독성이 높고 잔류기간이 길어 인체에 유해하고 환경 오염과 생태계 교란 문제가 점차 심화되고 있는 것으로 보고되었으며(Börner, 2009; RÖMPP Online, 2015), 오 · 남용하면서 약제 저항성을 획득한 해충과 선충들이 발생하여 방제효과 감소하는 등 여러 가지 문제점이 제기되고 있다(Kupferschmied 등, 2013). 최근에는 이러한 유기 합성 농약으로 인한 피해들이 증가하면서 해당 약제에 대한 사용이 금지되거나 제한되었고(Glare 등, 2012), 소비자들의 농약 사용에 대한 부정적 인식이 증가함에 따라 기존 방제 방법의 대안으로 생물학적 방제 등을 이용한 친환경 방제법 개발에 관심이 집중되고 있다(Stehle과 Schulz, 2015; Wachira 등, 2009). 식물병원성 해충과 선충을 친환경적으로 방제하기 위하여 진균, 세균, 곤충, 포식성 선충 등을 이용한 생물학적 방제법에 대한 연구가 활발히 이루어지고 있다. 최근 연구들에 따르면, 식물병원성 해충과 선충의 생물학적 방제법은 해충 및 선충의 행동 패턴을 변화시키는 페로몬과 같은 생물 유래 물질이나, 기생동물 또는 포식동물, 병원균, 진균, 바이러스 등의 생물체와의 상호작용을 이용하는 것으로 나누어 볼 수 있다(Bhattacharyya와 Jha, 2012).
곤충병원성 세균에 의한 해충의 생물학적 방제는 Bacillus thuringiensis를 이용하면서 시작되었는데, 1901년 병든 누에나방(Bombyx mori)으로부터 Bacillus sp.를 분리함으로써 Bacillus속 미생물에 의한 살충능력이 처음 밝혀졌다. 이후 이러한 그람-양성 세균에 의한 살충 효과 연구와 적용이 진행되었고, 최근에는 Pseudomonas, Photorhabdus, Xenorhabdus속 등 다양한 곤충병원성 세균들의 살충 활성과 병원성 인자에 대한 연구가 활발히 진행되고 있다. 또한 현재 750종의 진균이 곤충에 대한 병원성을 가지는 것으로 알려져 있는데, 진균은 곤충의 큐티클을 직접 침투하여 살충 효과를 내고(Fang 등, 2012; Gonzalez 등, 2016; Ramanujam 등, 2014; Wang 등, 2012), 살충 작용 메커니즘이 화학적 살충제에 비해 다양하기 때문에 곤충이 내성을 가지기가 힘들다는 장점이 있다(Kergunteuil 등, 2016). 해충 방제에 사용되는 생물제제 중에는 바이러스도 포함되는데 650여 개의 곤충병원성 바이러스가 보고되었으며, 곤충병원성 바이러스의 경우 기주 특이성이 높아 원하는 곤충만 선택적으로 방제할 수 있다는 장점이 있다(Kergunteuil 등, 2016; Moscardi 등, 2011; Ramanujam, 2014).
친환경 선충 방제제를 개발하기 위해 선충의 천적 미생물을 이용하는 방법도 주목 받고 있다. 선충의 천적 미생물은 선충포식성 진균과 선충병원성 세균이 있는데 이들 미생물은 선충 표면에 부착하여 표면을 분해하고 내부로 침투한 뒤 장을 파열시킨다. B. thuringiensis가 나타내는 살충 활성이 알려지고(Schnepf 등, 1998) 생물학적 살충제 연구가 시작된 후 살선충 활성을 나타내는 미생물을 탐색하는 연구들에서 다른 Bacillus종이나 Pseudomonas, Pasteuria속 세균 등 다양한 메커니즘으로 살선충 활성을 나타내는 세균들이 보고되었다(Tian 등, 2007). 일반적으로 널리 알려진 선충포식성 진균은 Arthrobotrys, Drechslerella, Dactylellina속으로 포획기관을 만들어 높은 살선충 활성을 나타내는 것으로 보고되었다(Yu 등, 2014). 다양한 방식으로 선충을 방제하는 미생물들의 병원성 인자와 메커니즘을 규명하기 위한 연구가 진행되고 있다.
본 리뷰에서는 최근까지 발표된 생물학적 방제제로 쓰일 수 있는 다양한 미생물들을 살펴보고, 이들이 해충과 선충을 치사시키는 메커니즘과 병원성 인자에 대하여 소개하려고 한다.
해충의 생물적 방제 방법
세균에 의한 해충의 생물적 방제 방법
해충을 방제하기 위해 생물학적 방제제로 가장 많이 사용되어 온 미생물은 B. thuringiensis로, Cry 독소라는 내독소를 생산하여 주로 나비목(Lepidoptera), 딱정벌레목(Coleoptera) 및 파리목(Diptera) 해충의 살충에 사용된다(Schnepf 등, 1998). B. thuringiensis는 획기적인 효과로 지금까지 전 세계적으로 사용이 확대되어 왔지만, 작물에 적용한 후 태양열이나 식물에서 나오는 다양한 화학 성분 등에 의해서 세균과 독소의 안정성이 감소하는 단점이 있다(Bravo 등, 2011). 또한 B. thuringiensis를 재배지에 반복적으로 사용했을 때 해충이 B. thuringiensis의 Cry 독소에 대한 내성이 보고되면서 B. thuringiensis를 대체하여 생물학적 방제제로 이용할 수 있는 미생물이 요구되고 있다(Bravo 등, 2011).
최근 미생물을 이용한 해충 방제에 있어 Pseudomonas속이 각광을 받고 있는데, 지난 50여 년 동안의 연구에서 Pseudomonas속은 식물 근권에서 우점하며 기주 식물의 생장을 촉진시키고, 병원성 미생물과 진균으로부터 식물을 보호하여 식물의 저항성을 증가시키는 등의 유익한 활성을 가지는 것으로 보고되었다(Haas와 Défago, 2005). 최근에는 이 속의 비교 유전체 분석을 통하여 곤충에 대한 독소를 생성하는 유전자가 밝혀지면서 지금까지 몰랐던 해충 방제 능력에 대한 연구가 활발히 진행되고 있다. 따라서 본 단락에서는 Pseudomonas속 미생물을 포함하는 B. thuringiensis를 대신하여 생물학적 방제에 사용할 수 있는 미생물에 대해 논의하고자 한다.
① Pseudomonad
Gamma proteobacteria에 속하는 그람-음성 세균인 Pseudomonas속은 바다, 토양, 식물체 표면 및 곤충의 장내에까지 지구상의 다양한 장소에서 분리되었다(Silby 등, 2011). 그 중 P. protegens 종은 식물 근권에 서식하면서 식물체를 보호하는 것으로 알려져 있다(Haas와 Défago, 2005). 대표적으로 알려져 있는 P. protegens CHA0와 Pf-5를 살펴보면, 먼저 P. protegens CHA0는 1986년 스위스의 담배 뿌리에서 처음으로 동정되었고, P. protegens Pf-5는 미국 밀밭에서 발견되었는데 이 두 가지 균주들은 주로 식물 병원성 진균 방제에 대한 다양한 메커니즘들이 보고되었다(Mark 등, 2006). 최근 유전체 분석을 통해 살충 효과에 관여하는 fit (fluorescence insecticidal toxin) 유전자가 발견되면서 진균 방제 이외에 식물 병원성 해충의 생물학적 방제 가능성이 제시되고 있다(Péchy-Tarr 등, 2008). Pseudomonas spp.는 식물의 근권에 서식하기 때문에 토양 유래의 곤충에 대한 접촉이 용이하고 지상부에 비해 태양열이나 건조에 대한 생존율이 증가할 수 있다. P. protegens뿐만 아니라 새롭게 재분류된 P. fluorescens, P. chlororaphis, P. entomophila와 P. taiwanensis 등이 해충에 대한 살충 활성을 나타낸다고 보고되었다(Flury 등, 2016; Olcott 등, 2010; Péchy-Tarr 등, 2008; Ruffner 등, 2015). 이 단락에서는 Pseudomonas속 세균이 곤충에 살충 활성을 나타내는 작용메커니즘과 병원성 인자로 작용하는 독소에 대해서 알아보고자 한다(Fig. 1A, 2A, Table 1).
Fig. 1
Insecticidal capacity of the pathogenic bacteria. (A) Pseudomonas spp. secrete insecticidal toxins and enzymes such as rhizoxin, orfamide, O-antigenic polysaccharide, Fit (fluorescence insecticidal toxin) toxin, chitinase, and protease. (B) Cry proteins produced by Bacillus spp. are activated to δ-toxins and toxins only in the insect gut for killing insect pest. (C) Photorhabdus spp. and Xenorhabdus spp. that secrete insecticidal toxins, Mcf, Tcs, Pir, and PVC invade insect larva during entomopathogenic nematode infection.
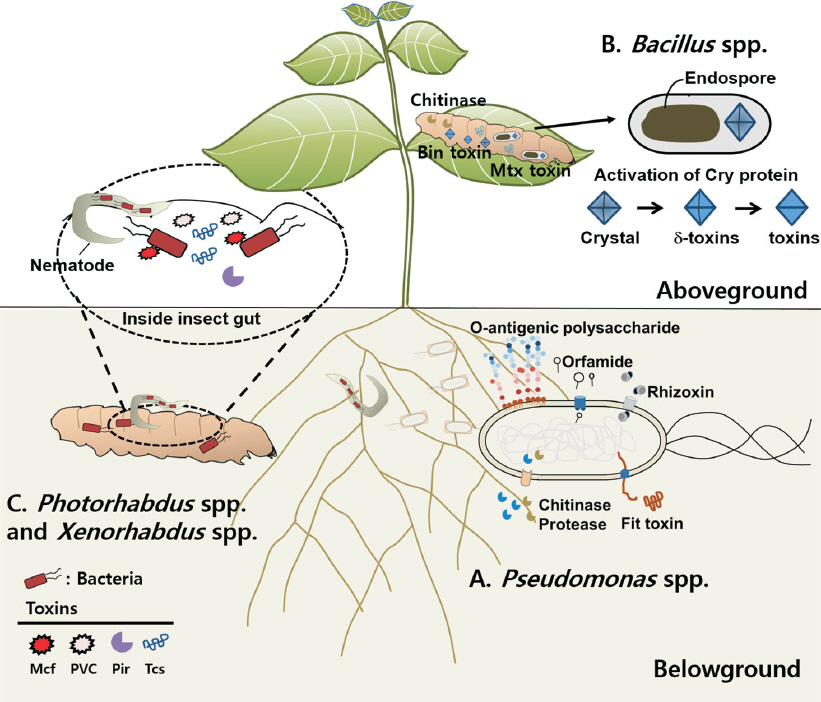
Fig. 2
Chemical structure of insecticidal and nematicidal determinants derived from bacteria and fungi. Insecticidal microbial determinants (A) and nematicidal microbial determinants (B). The broken line inset box indicates the common microbial determinants of both insecticidal and nematicide capacity.
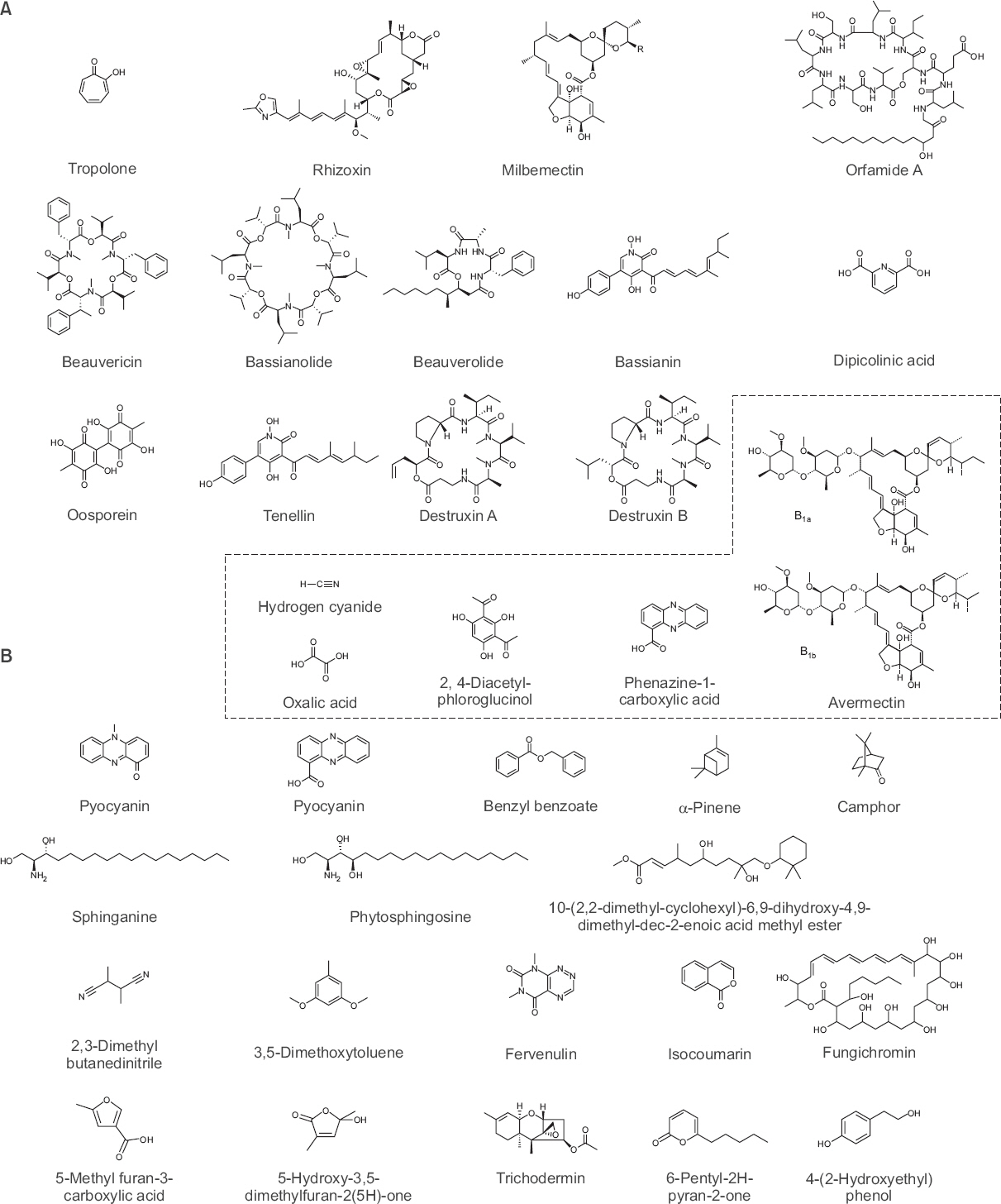
Table 1
Microbes, their microbial determinants, and the commercial products for controlling the insect pests
| Species | Target insects | Effector/determinant | Products | References | |
|---|---|---|---|---|---|
| Bacteria | |||||
| Pseudomonas fluorescens | Aphidoidea spp.; Isoptera spp.; Epilachna vigintioctopunctata; Cnaphalocrocis medinalis | Lipopeptides; Orfamide; Chitinase; 2,4-Diacetylphloroglucinol; Hydrogen cyanide | Bio-pseudo, Monas | Devi and Kothamasi, 2009; Hashimoto, 2002; Jang et al., 2013 | |
| P. protegens | Lepidopteran spp.; Dipteran spp.; Drosophila melanogaster | Fit; Rizoxin; Orfamide A; GacA; Chitinase; Phospholipase | Flury et al., 2016; Loper et al., 2016; Péchy-Tarr et al., 2008; Ruffner et al., 2015 | ||
| P. chlororaphis | Lepidopteran spp. | Fit; O-PS; Chitinase | Kupferschmied et al., 2016; Péchy-Tarr et al., 2008 | ||
| P. entomophila | D. melanogaster | Protease AprA and AprX | Liehl et al., 2006 | ||
| P. taiwanensis | D. melanogaster | tccC; Tc-related gene | Liu et al., 2010 | ||
| Lysinibacillus sphaericus | Culex spp.; Anopheles spp.; Aedes spp.; Simuliidae spp.; Nematocera spp. | Bin; Mtx | VectoLex | Charles et al., 2000; Ruiu, 2015 | |
| Paenibacillus sp. D1 | Helicoverpa armigera | Chitinase | Singh et al., 2016 | ||
| P. lentimorbus P. popilliae Paenibacillus spp. Kh3 | Coleoptera spp. | Cry | Milky Spore | Gorashi et al., 2014; Zhang et al., 1997 | |
| Brevibacillus laterosporus | Coleoptera spp.; Diptera spp.; Plutella xylostella; Leptinotarsa decemlineata | Chitinase ChiA1 | Prasanna et al., 2013 | ||
| Photoharbdus luminescens P. asymbiotica | Manduca sexta; L. decemlineata; Bemisia tabaci | Mcf; Pir1; Tcs; PVC | Bowen and Ensign, 1998; Daborn et al., 2002; ffrench-Constant et al., 2007; Waterfield et al., 2005; | ||
| Xenorhabdus nematophila | P. xylostella | Tcs; Phenethylamide compounds; MrxA; XaxAB | Nepuwang, Dipeukil, Jayeonilang-Nepeukil | Bowen and Ensign, 1998; Jung and Kim, 2006 | |
| Serratia entomophila S. proteamaculans | Costelytra zealandica, | Sep protein | Carnosine, Serrapeptase | Hurst et al., 2000; Nuñez-Valdez et al., 2008 | |
| S. marcescens | Orthoptera spp.; Coleoptera spp.; Hymenoptera spp.; Lepidoptera spp.; Diptera spp.; Rhagoletis pomonella | Metalloprotease | Ishii et al., 2014; Kwak et al., 2014 | ||
| S. nematodiphila | H. armigera; Spodoptera litura | Chitinase; Lipase; Protease | Kwak et al., 2015; Patil et al., 2013 | ||
| Burkholderia plantarii | Dermatophagoides spp.; Tyrophagus putrescentiae; Coptotermes formosanus | Tropolone | Kil et al., 2014; Morita et al., 2003 | ||
| B. rinojensis | Spodoptera exigua; Tetranychus urticae | Unknown (secondary metabolite and heat stable) | Cordova-Kreylos et al., 2013; Takeshita et al., 2015 | ||
| Chromobactierum subtsugae | P. xylostella; B. tabaci; Nezara viridula; Diabrotica undecimpunctata; Diabrotica virgifera; L. decemlineata; Aethina tumida | Unknown (heat stable) | Grandevo® | Martin et al., 2004, 2007a, 2007b | |
| Streptomyces phaeochromogenes | D. melanogaster; Spodoptera littoralis | Avermectins | Yuan-Mei Streptomyces spp, Cheonggotan | Bream et al., 2001; Pitterna et al., 2009 | |
| Fungi | |||||
| Beauveri bassiana | Aphidoidea spp.; Thysanoptera spp.; Psyllidae spp.; Curculionoidea spp.; Pseudococcidae spp.; Trialeurodes vaporariorum; P. xylostella; Galleria mellonella | Beauvericin; Bassianin; Bassianolide; Beauverolides; Beauveriolides; Tenellin; Oosporein; Oxalic acid; Bassiacridin; Insect toxin protein Bb70p | Naturalis, Botanigard, BotaniGard, Mycotrol, balance, CornGard, Racer, Bb-Protec, Bae-Sin, Bassianil, PHC BEA TRON, Conidia, Ostrinol, Myco-Jaal, Biosoft, Biowonder, Nuturalis-L, Seremoni, Tory, Seonchungae | Duarte et al., 2016; Gonzalez et al., 2016; Khan et al., 2016; Malekan et al., 2015; Mascarin and Jaronski, 2016; Quesada-Moraga et al., 2006; Ramanujam et al., 2014; Safavi, 2013; Xu et al., 2009 | |
| Metarhizium anisopliae | Anisoplia austriac; Cleonus punctiventris; Otiorhynchus ovatus; Tetranychus evansi; Megalurothrips sjostedti | Destruxins A, B, C, D, E, F | Taerain, Met52EC, BIO 1020, Bio-BlastTM, Bio Magic, Multiplex, Metarhizium | Gonzalez et al., 2016; Kershaw et al., 1999; Maniania et al., 2016; Mfuti et al., 2017; Ramanujam et al., 2014; Roberts and St. Leger, 2004 | |
| Lecanicillium muscarium | B. tabaci; Rhynchota spp.; Ricaniidae spp.; T. vaporariorum | Bassianolide; Dipicolinic acid | Mycotal, Vertalec, Inovert, Biocatch, Verticare, Rhampid | Cuthbertson et al., 2008; Duarte et al., 2016; Gonzalez et al., 2016; Güçlü et al., 2010; Jirakkakul et al., 2008; Malekan et al., 2015; Ramanujam et al., 2014 | |
| Isaria spp. | T. vaporariorum; Diaphorina citri; Toxoptera citricida; Homalodisca vitripennis; Diaprepes abbreviatus | Chitinase gene Ifchit1 | PreFeRal, Nofly, PFR-97TM, Prioroty, Sunchungzero, Myeoldie | Avery et al., 2016; Gonzalez et al., 2016; Huang et al., 2016; Hunter et al., 2011; Ramanujam et al., 2014; Zimmermann, 2008 | |
| Virus | |||||
| Baculovirus | |||||
| NPV | AcMNPV | Autographa californica; S. exigua; Hellulo undalis; Pectinophora gorrypiella | VP39; P78; VP80; Arif-1; Ac004; Ac102; Ac152; P78/83; EXON0; Ac93; EC27; Lefs; PK1; P35; P49 | VPN-ULTRA | Haase et al., 2015; Monteiro et al., 2012; Sun, 2015 |
| HearNPV | H. armigera; Heliothis assulta | VP39 | Helicovex, Diplomata, Heli-Cide®, Heliokill® | Gonzalez et al., 2016; Haase et al., 2015; Monteiro et al., 2012; Ramanujam et al., 2014; Yang et al., 2012 | |
| HezeSNPV | Helicoverpa zea | Gemstar, HzNPV CCAB | Haase et al., 2015; Lacey et al., 2015; Rowley et al., 2011 | ||
| MabrNPV | Mamestra brassicae; H. armigera; S. exigua | Mamestrin | Sun, 2015 | ||
| OrpsNPV | Orygia pseudotsugata | IAPs | TM Biocontrol | Monteiro et al., 2012; Moscardi et al., 2011 | |
| SpliNPV | Spodoptera littoralis | P35 | Littovir, Spodo-Cide®, Spodopterin® | Gonzalez et al., 2016; Monteiro et al., 2012; Ramanujam et al., 2014 | |
| GV | AdorGV | Adoxophyes orana | Capex | Lacey et al., 2015; Nakai, 2009 | |
| CpGV | Cydia pomonella | IAPs | Cyd-X, Virosoft, Carpovirusine, Carpovirus Plus, Capex, Granupon, Virin Cyap, Madex, Madex Twin | Arthurs and Lacey, 2004; Haase et al., 2015; Monteiro et al., 2012 | |
| CrleGV | Cryptophlebia leucotreta | Cryptogran, Cryptex | Haase et al., 2015; Moore et al., 2015 | ||
| PhopGV | Phthorimaea operculella | Baculovirus Corpoica, PTM baculovirus, Matapol Plus, Bacu-Turin | Arthurs et al., 2008; Haase et al., 2015; Lacey et al., 2015 | ||
| PiraGV | Pieris rapae | Yang et al., 2012 | |||
| PlxyGV | P. xylostella | Yang et al., 2012 | |||
| Reoviridae | |||||
| Cypovirus | Dendrolimus punctatus; D. kikuchii; D. spectabilis; D. supreans | - | - | Lacey et al., 2015; Yang et al., 2012 | |
| Nudiviruses | |||||
| Oryctes virus | Oryctes rhinoceros | - | - | Huger, 2005; Lacey et al., 2015; Ramle et al., 2005 | |
Fit 독소(fluorescence insecticidal toxin): P. protegens와 P. chlororaphids의 Fit 독소는 8개의 유전자(fitABCDEFGH) 클러스터로 구성되어 있으며 곤충에게 특이적 독소로 작용한다. 그 중, fitA, fitB, fitC 및 fitE는 type I secretion system을 발현시켜 Fit 독소가 세포 밖으로 분비될 수 있도록 하며, fitF, fitG와 fitH는 곤충의 장내환경을 인식해 Fit 독소가 곤충 장내에만 발현되도록 한다(Kupferschmied 등, 2014). 또한, fitD는 곤충의 장내에서 세포사멸(apoptosis)을 유도하여 장세포를 파열함으로써 살충 활성을 보인다(Kupferschmied 등, 2013). 하지만 P. protegens와 P. chlororaphids종의 유전체에서 fit 유전자를 제거하면 살충 활성이 감소하지만 여전히 살충 활성을 나타내는 것으로 미루어 Pseudomonas spp.에는 Fit 독소 이외의 다른 병원성 인자 또한 부분적으로 살충 활성에 작용하는 것으로 추정된다(Péchy-Tarr 등, 2008; Ruffner 등, 2013).
Rhizoxin: Rhizoxin은 이전에 식물병원성 진균인 Rhizopus microsporus로부터 분리된 이차 대사산물로 알려져 있으며 초파리에 살충 활성을 나타내는 병원성 인자로 보고되었다(Loper 등, 2016). 최근 식물 근권 세균인 P. protegens Pf-5에서도 rhizoxin이 발견되었는데, 유전체상의 rhizoxin은 fit 유전자와 마찬가지로 유전체의 외래유입 유전자 부위에 위치하였다. 이는 P. protegense가 수평적 유전자 이동(horizontal gene transfer)에 의해 유전자를 획득했을 것으로 추정하고 있다(Loper 등, 2016; Ruffner 등, 2015). Rhizoxin은 16개의 매크로라이드(macrolide)의 구조를 가지는 항생물질로, 진핵생물의 유사분열 과정 중에 베타튜블린(β-tubulin)과 결합하여 미세관(microbutule) 형성을 방해하는 독소로 작용한다(Gupta와 Bhattacharyya, 2003).
Orfamide A: Pseudomonas spp.의 이차 대사산물 중 지질 펩타이드의 일종인 orfamide A는 초파리 및 진딧물(Myzus persicae) 유충에 대해 살충 활성을 나타내는 것으로 보고되었다(Jang 등, 2013; Loper 등, 2016; Ma 등, 2016). Orfamide는 주로 곤충의 구강으로 침투하여 경구 독성을 나타내는 것으로 알려져 있다(Gross와 Loper, 2009). 현재까지 Pseudomonas속 세균들에서 orfamide A와 유사한 orfamide B-G가 발견되었으며 orfamide B는 식물병원균으로 알려진 Rhizoctonia solani에 대해 항진균 활성도를 나타내는 것으로 보고되었다(Ma 등, 2016). Orfamide B-G의 해충에 대한 살충 활성에 대해서는 아직 자세히 알려지지 않았지만 orfamide A와 유전자의 염기서열이 유사하기 때문에 해충에 대한 살충 활성을 나타낼 것으로 기대한다.
O-antigenic polysaccharide (O-PS): 그람-음성 세균인 Pseudomonas spp.의 외부 세포막은 지질다당류(lipopolysaccharide)의 구성성분인 lipid A와 core oligosaccharide 및 O-antigenic polysaccharide (O-PS) 등으로 구성되어 있다(Lam 등, 2011; Raetz와 Whitfield, 2002; Trent 등, 2006). 세포막을 구성하고 있는 O-PS는 숙주-미생물 간 상호작용에서 숙주의 방어 기전을 억제하는 중요한 병원성 인자인 것으로 보고되었다(Lam 등, 2011; Lerouge와 Vanderleyden, 2002). Pseudomonas spp.의 유전체 분석을 통하여 식물유용균으로 알려진 P. protegens CHA0와 P. chlororaphis PCL1391의 O-PS 합성에 관련된 유전자군(O-specific antigen, OSA)이 인체병원균인 P. aeruginosa와 다른 새로운 유전자군을 획득한 것으로 나타났는데 이는 인체뿐만 아니라 곤충과 선충 등 다양한 숙주를 감염시키기 위해 진화했을 것으로 추정되었다. P. protegens CHA0의 OSA 돌연변이주를 모델 곤충인 꿀벌부채명나방(Galleria mellonella)에 접종하였을 때 살충 활성이 감소하는 것으로 보고되어(Kupferschmied 등, 2016), Pseudomonas spp.의 살충 활성에 있어 O-PS가 곤충의 방어 메커니즘을 억제하는 중요한 병원성 인자라고 생각된다.
키틴분해효소: 해충 장내로 침입한 Pseudomonas spp.는 키틴으로 구성된 해충 장벽에 도달하게 된다. Pseudomonas spp.는 장내에서 키틴분해효소를 분비하여 해충의 장벽을 분해하고 병원성을 띠게 된다. 유전체 분석을 통해 P. protegens, P. fluorescens, P. chlororaphis, P. pisciumd와 P. aureofaciens 및 다양한 Pseudomonas spp.의 유전체에서 키틴분해효소가 발견되었다(Flury 등, 2016). 특히, P. protegens CHA0에서 키틴분해효소인 chiC를 돌연변이시켰을 때, 배추좀나방 유충에 대한 살충 활성이 감소한 것으로 나타나 키틴분해효소가 살충 활성에 중요한 병원성 인자인 것으로 확인되었다(Flury 등, 2016).
단백질분해효소: 해충은 침입한 병원균에 대한 면역반응의 일종으로 항균 펩타이드를 생산하는데, 병원균은 serralysin (Zn-dependent metallopeptidase AprX [alkaline protease X])과 염기성 단백질분해효소 AprA (alkaline protease A)와 같은 단백질분해효소를 생산함으로써 곤충의 면역 반응을 억제시킨다. AprA와 AprX는 곤충의 조직을 손상시키고 과민반응(anaphylactic)을 일으키게 한다(Park과 Ming, 2002). 특히 aprA를 돌연변이시킨 P. entomophila에서 D. melanogaster에 대한 병원성이 감소하는 것으로 나타나, 염기성 단백질분해효소에 의한 병원성이 검증되었다(Liehl 등, 2006).
② Bacilli
Lysinibacillus sphaericus: 캘리포니아의 모기(Culiseta incidens) 유충에서 처음 분리되었으며, 그람-양성균으로 포자를 형성하는 세균이다. L. sphaericus는 B. thuringiensis처럼 Cry 독소와 Bin 독소(Binary prototoxin)라는 살충 활성을 나타내는 독소를 생성한다(Fig. 1B; de Oliveira 등, 2004). Bin 독소는 소체 결정(parasporal crystal) 안에 42 kDa의 BinA와 51 kDa의 BinB 단백질로 구성되어 있다. 곤충이 이 독소들을 섭취하게 되면 장에서 용해되어 저분자 유도체로 분해된 후 곤충의 중장 상피세포에 손상을 주어 살충 작용을 한다(Charles 등, 2000). 또한 L. sphaericus는 영양생장기에 Mtx 단백질(mosquitocidal toxins)을 생성하는 것으로 보고되었지만 자세한 기전에 대해서는 알려지지 않았다. L. sphaericus는 모기, 진딧물, 혹파리(Thecodiplosis japonensis)와 깔따구(Chironomus plumosus) 등에 살충 활성을 나타내는 것으로 알려져 있어 다양한 해충에 사용 가능한 생물학적 살충제로 개발할 수 있을 것이다(Fig. 1B).
Paenibacillus spp.: 1872년 최초로 분리되었을 때에는 막대기 모양의 내생포자를 형성하는 호기성(혹은 조건적 혐기성) 그람-양성 세균의 특징을 토대로 Bacillus속에 속하였으나 1993년에 염기서열 분석 및 계통학적 분석을 통하여 Paenibacillus속으로 분류체계가 정정되었다(Ash 등, 1993). Paenibacillus속 미생물은 사람, 동물, 식물체 등의 다양한 환경에서 발견되는데, 대부분은 토양에서 발견되며 식물의 근권에 서식하면서 식물의 생장을 촉진시키는 역할을 하는 것으로 알려져 있다. 또한 Paenibacillus larvae와 P. popilliae는 벌목(Hymenoptera), 딱정벌레목, 나비목 곤충에 대해 병원성을 나타내는 것으로 보고되었다(Neung 등, 2014; Sharma 등, 2013). Paenibacillus spp.의 대표적인 병원성 인자는 키틴분해효소와 Cry 독소이다. 키틴분해효소는 곤충의 외골격과 장 내벽을 이루는 키틴을 분해하는 것으로 알려져 있으며, Paenibacillus sp. D1이 생성하는 키틴분해효소는 왕담배나방(Helicoverpa armigera)에 대해 살충 활성을 나타내는 것으로 보고되었다(Singh 등, 2016). P. lentimorbus는 B. thuringiensis에서 잘 알려진 Cry 독소와 유사한 Cry 독소를 만들어 딱정벌레에 대해 살충 활성을 보이는 것으로 보고되었다. 그 밖에도, P. popilliae와 Paenibacillus sp. Kh3 등이 Cry 독소를 생성하는 것으로 알려져 있다(Fig. 1B; Gorashi 등, 2014; Zhang 등, 1997).
Brevibacillus laterosporus: B. laterosporus는 토양, 물, 곤충의 몸 속, 식물체 표면 등 다양한 곳에서 분리되는데, 막대기 모양의 내생포자를 형성하는 그람-양성 세균으로 canoe-shaped parasporal body를 생성하는 것이 특징이다(Ruiu, 2013). 1916년 Laubach에 의해 담수에서 분리되어 B. laterosporus로 동정되었으나 16S rRNA 염기서열 분석을 통해 Brevibacillus속의 Brevibacillus laterosporus로 정정되었다(Shida 등, 1996). B. laterosporus는 나비목, 딱정벌레목과 파리목에 대하여 병원성을 나타내는 것으로 보고되었다(Ruiu, 2013). B. laterosporus의 키틴분해효소인 ChiA1이 배추좀나방에 대한 살충 활성을 나타낸다고 알려졌다(Prasanna 등, 2013). Brevibacillus속 미생물의 살충 메커니즘은 잘 알려져 있지 않아 추가적인 병원성 인자의 특성 및 메커니즘 연구가 필요하다(Fig. 1B).
③ Photorhabdus spp.와 Xenorhabdus spp
Enterobacteriaceae에 속하는 Photorhabdus spp.와 Xenorhabdus spp.는 각각 곤충병원성 선충인 Heterorhabditidae와 Steinernematidae의 장 내부에 공생하며 살충 활성을 나타내는 세균이다. 살충 작용을 위하여 곤충의 구강으로 침투해야 하는 B. thuringiensis나 Pseudomonas spp.와 다르게 Photorhabdus sp.와 Xenorhabdus sp.는 토양에 서식하는 곤충병원성 선충과 공생에 의해 기주에 감염한다(Fig. 1C; Lacey와 Georgis, 2012). 기주 내부로 침입한 선충공생균들은 항균 화합물, 분해효소와 같은 병원성 인자를 생성하여 곤충을 치사시킨다(Table 1; Ruiu, 2015). 특히 Photorhabdus sp.와 Xenorhabdus sp.가 가지고 있는 Mcf (makes caterpillars floppy) 유전자는 살충 활성을 나타내는 중요한 병원성 인자로 알려져 있다(Lacey와 Georgis, 2012). Photorhabdus sp.는 곤충의 면역체계에 의해 만들어지는 항세균 펩타이드에 대항하기 위해 지질다당류(lipopolysaccharide)를 생산한다. X. nematophila는 곤충 체내에서 증식하면서 곤충이 만들어내는 항균 펩타이드의 발현을 방해하여 세균 자체를 보호하는 것으로 알려져 있다. 본 단락에서는 Photorhabdus spp.와 Xenorhabdus spp.가 생산하는 독소에 대하여 알아보겠다.
Mcf (makes caterpillars floppy) 독소: Photorhabdus luminescens W14에서 처음으로 분리된 Mcf1 독소는 담배를 기주로 하는 박각시나방(Manduca sexta) 애벌레에 대해 살충 활성을 나타내는 것으로 보고되었다(Daborn 등, 2002). Mcf1 독소는 세포예정사(apoptosis)에 관여하는 BH-3 도메인을 통해 곤충의 중장 상피(midgut epithelium)와 혈액세포(hemocyte)를 빠르게 파괴하여 살충 활성을 나타내는 것으로 알려져 있다(Dowling 등, 2007). 이후 P. asymbiotica, Xenorhabdus nematophila 및 Xenorhabdus bovienii에서도 살충 활성을 나타내는 Mcf와 유사한 독소가 발견되었다(Sugar 등, 2012; Wilkinson 등, 2009). P. luminescens의 Mcf1, Mcf2 독소는 Pseudomonas속 세균의 Fit 독소와 유사한 것으로 보고되었는데, 이는 수평적 유전자 이동(horizontal gene transfer)에 의해 동일한 조상으로부터 mcf와 fit 유전자가 유래하였을 것으로 추정된다(Ruffner 등, 2015).
Pir (Photorhabdus insect related) 독소: P. luminescens는 Pir 단백질을 생성하는 것으로 알려져 있다(Lacey 등, 2015). PirA와 PirB로 구성된 Pir 단백질은 유충의 발달을 방해하는 유충 호르몬(juvenile hormone esterase)이다. 특히 PirB는 B. thuringiensis의 Cry 독소와 유사하다고 알려져 있어 Cry 독소의 대안으로서 PirB 단백질이 주목 받고 있으며, 실제로 PirB 유전자를 이용한 재조합 작물을 제작하였을 때 해충에 대한 저항성이 증대되는 것으로 보고되었다(Waterfield 등, 2005).
Tcs (toxin complexes) 독소: Tcs는 Photorhabdus spp. 유전체의 PAI1 (pathogenicity island 1)에서 발현되는 tca, tcb, tcc와 tcd로 이루어져 있으며 살충 활성을 나타내는 독소이다. P. luminescens W14에서 최초로 분리된 Tcs 독소는 박각시나방에 대해 살충 활성을 나타냈으며 P. luminescens W14의 살충 활성과 관련이 있는 것으로 밝혀졌고, X. nematophila에서도 Tcs 독소와 유사한 독소가 발견되었다(Bowen과 Ensign, 1998). 또한 Photorhabdus sp.와 Xenorhabdus sp.의 ABC complex도 tcs와 유사한 유전자로 알려졌는데, 곤충의 구강을 통하거나 직접 접종 시 살충 활성을 나타내는 것으로 보고되었다. ABC complex는 A, B와 C의 고분자 대단위체로 이루어져 있으며, 살충 활성에 중요한 병원성 인자이다(ffrench-Constant와 Waterfield, 2005). Photorhabdus sp.와 Xenorhabdus sp.뿐만 아니라 Serratia entomophila, Yersinia sp., Paenibacillus sp. 등의 곤충병원성 세균에서도 ABC complex가 존재하는 것으로 보고되어 ABC complex는 박각시나방 등의 해충을 방제하기 위한 생물학적 방제제로 개발하여 이용이 가능할 것으로 생각된다.
PVC (Photorhabdus virulence cassettes): Photorhabdus sp.의 PVC 독소는 S. entomophila와의 상동성 분석을 통해서 규명되었으며 P. luminescens의 Mcf 독소, 인체 병원균 Clostridium difficile의 toxin A와 염기서열이 유사한 것으로 알려져 있다(ffrench-Constant 등, 2007). PVC에 의해 생성된 단백질은 직접적인 항균 활성은 없지만 곤충의 혈액세포(hemocyes)를 파괴할 수 있다. P. asymbiotica와 P. luminescens의 PVC에 의해서 꿀벌부채명나방에 대한 살충 활성이 보고되었지만 기주 곤충에 대한 실증실험이 이루어지지 않아 추가적인 연구가 필요하다.
④ Serratia spp
Gamma proteobacteria에 속하는 그람-음성 세균인 Serratia는 물, 토양, 동물 및 식물의 표면에서 분리되었다(Grimont와 Grimont, 1992). Serratia속 미생물은 항균 화합물을 포함하여 다양한 이차 대사산물을 생성하는 것으로 알려져 있는데(Harris 등, 2004), 식물생장촉진 화합물, 항진균성 화합물을 생산하고, 질소고정 공생 미생물들의 군집화를 유도하며 기주 곤충에 따라 공생 세균과 병원균으로 분류된다(Kalbe 등, 1996; Zhang 등, 1996). Serratia spp.는 종에 따라서 다양한 병원성 인자들을 생성하는데 Xenorhabdus spp., Photorhabdus spp.와 유사한 병원성 인자가 보고되었다(Hurst 등, 2000).
S. entomophila와 S. proteamaculans는 뉴질랜드풍뎅이(Costelytra zealandica)에 황색병(amber disease)을 야기하는 것으로 알려져 있으며(Jackson 등, 2001), Sep 단백질(Serratia entomophila pathogenicity)이 병원성 인자로 알려져 있는데 앞서 언급한 P. luminescens의 Tcs 독소와 유사하다(Hurst 등, 2000). S. marcescens HR-3에서도 살충 활성을 나타내는 단백질이 분리되었는데, 아미노 말단 염기서열이 S. marcescens SM6, E15의 금속단백질 분해효소와 유사하며 61 kDa에 달하는 단량체 형태이고 단백질을 분해하는 것으로 보고되었다(Tao 등, 2006). 이후, S. marcescens Bizio가 분비하는 serralysin의 금속단백질 분해효소가 곤충 면역감시 세포의 점착 능력을 감소시켜 면역세포의 기능을 억제하는 것으로 나타나 금속단백질 분해효소에 의한 살충 활성이 검증되었다(Ishii 등, 2014). 또한 곤충병원성 선충(Heterorhabditidoides chongmingensis)에 기생하는 S. nematodiphila DSM 21420T는 유전체 염기서열 분석을 통해 단백질분해효소, 지질분해효소, 키틴분해효소 등 가수 분해적 특징을 가진 살충 활성인자들을 생성하는 것으로 확인되었다(Kwak 등, 2015).
⑤ Burkholderia spp
Burkholderia속 미생물은 Betaproteobacteria에 속하는 그람-음성 세균으로 물, 토양, 곤충, 진균 및 식물의 근권 등 다양한 환경에서 서식할 수 있다. B. cepacia의 경우 양파에 병을 유발하는 병원균으로 알려져 있지만(Burkholder, 1950), 반대로 Burkholderia속의 다른 종은 식물 병원균에 대하여 식물에 유익한 역할을 하는 것으로 알려져 있다. 또한 Burkholderia spp.는 키틴분해효소와 단백질분해효소를 생성하는데 곤충의 큐티클과 내장을 분해하여 살충 활성을 나타내는 것으로 보고되었다(Bunnori와 Mohamed, 2012; Kong 등, 2001). 추가적으로 Burkholderia spp.는 단백질 분해효소, 지질 분해효소, 용혈성 항체, 독소 등을 생성한다고 알려져 살충 활성에 대한 가능성이 제시되고 있다(Kil 등, 2014).
Burkholderia spp.는 비 벤제노이드 방향족 화합물인 트로폴론(tropolone, 2-hydroxy-2,4,6-cycloheptatrien-1-one)을 생산하는데 살충 활성과 함께 항세균성, 항진균성 활성을 나타내는 특징을 가지고 있다(Fig. 2A; Morita 등, 2003). 특히 B. plantarii에 의해 생성되는 트로폴론은 긴털가루진드기(Tyrophagus putrescentiae), 큰다리먼지진드기(Dermatophagoides farina), 대만지하흰개미(Coptotermes formosanus) 등에 대해 살충 활성을 나타내는 것으로 보고된 바 있다(Morita 등, 2003). B. cepacia의 경우 페놀을 포함하는 화합물을 생산하여 초파리 유충의 tyrosinase를 억제하고 살충 활성을 나타내는 것으로 보고되었다(Hsu 등, 2014). B. rinojensis A396은 일본의 토양에서 처음 분리되었는데 파밤나방(Spodoptera exigua)과 점박이응애(Tetranychus urticae)에 대하여 살충 활성을 나타내었다(Takeshita 등, 2015). 대부분의 곤충 병원균은 곤충이 섭취한 후 살충 활성을 나타내는 것으로 알려져 있는데 B. rinojensis A396은 섭취뿐만 아니라 접촉에 의해서도 살충 활성을 나타낸다고 보고되었다(Cordova-Kreylos 등, 2013). 또한 고온 처리한 배양액에서도 살충 활성을 나타내기 때문에 이차 대사산물이나 열에 안정한 효소가 살충 활성과 연관이 있을 것으로 생각된다. 이후 Burkholderia spp.의 병원성 인자와 살충 작용에 대한 기전을 밝히는 연구가 수행된다면 안정적인 생물학적 방제제로서 이용이 가능할 것이다(Cordova-Kreylos 등, 2013).
⑥ Chromobacterium spp
Beta proteobacteria에 속하는 그람-음성 세균인 Chromobacterium spp.는 일반적으로 토양이나 물에서 발견되었다(Hungria 등, 2005). 최초로 동정된 1880년부터 2000년 초반까지, Chromobacterium violaceum Bergonzini는 Chromobacterium을 대표하는 보랏빛 색소를 가진 세균이었다. 1880년 동정된 C. violaceum은 보라색 색소인 바이올라세인(violacein)을 생성하는 것으로 보고되었는데, 이것은 trypanocide로 항균성, 항암성의 활성을 나타내었다(Durán 등, 1994). 또한 이렇게 보랏빛 색을 띠는 미생물들은 콜로라도감자잎벌레의 유충과 여러 종의 해충에 대해서 구강을 통해 독성을 나타내는 것으로 알려져 있다(Martin 등, 2004).
C. violaceum ATCC 12472T는 유전체 염기서열 분석을 통해 살충성 유전자가 발견되었다(Brazilian National Genome Project Consortium, 2003). C. violaceum ATCC 12472T는 콜로라도감자잎벌레에 대한 살충 활성을 나타내지 않았지만 P. luminescens의 살충 단백질인 CV1887과 유사한 염기서열을 가지고 있는 것으로 밝혀졌다(Brazilian National Genome Project Consortium, 2003). 2007년도에는 미국의 독미나리(hemlock) 나무 토양에서 C. subtsugae가 처음 발견되었는데, 한천배지에서 배양 시 콜로니가 짙은 보라색을 보였고 콜로라도감자잎벌레에 대해 살충 활성을 나타내는 것으로 보고되었다(Martin 등, 2004). 또한, C. subtsugae는 배추좀나방, 담배가루이(Bemisia tabaci), 남쪽풀색노린재(Nezara viridula L.), 오이딱정벌레(Diabrotica undecimpunctata), 옥수수뿌리벌레(D. virgifera), 작은벌집딱정벌레(Aethina tumida)에 대해서도 살충능력을 나타내었다(Martin 등, 2007a, 2007b). C. subtsugae는 기주 곤충이 다양하고 살충 활성을 나타내는 독소가 열에 안정한 것으로 보고되었으며(Martin 등, 2007c), C. subtsugae의 추출물은 현재 미국환경보호국의 승인을 받아 Grandevo® 라는 이름의 유기인산 살충제로 상업화되었다(Marrone Bio Innovations, Davis, CA, USA).
C. violaceum과 C. subtsugae 외에도 C. aquaticum, C. haemolyticum, C. piscinae, C. pseudoviolaceum, C. vaccinii 및 C. amazonense가 발견되었지만 살충 활성에 대한 자세한 연구는 되어 있지 않다(Han 등, 2008; Kämpfer 등, 2009; Menezes 등, 2015; Soby 등, 2013; Young 등, 2008).
⑦ 방선균(Stretomycetes)
방선균은 Actinobacteria에 속하는 그람-양성 세균으로 다양한 환경에서 살 수 있는데, 진균과 유사한 사상 구조(filamentous)를 가지는 것이 특징이다. Streptomyces속의 가장 두드러진 특징은 항진균성, 항균성, 항바이러스성, 항암성, 살충성, 면역억제 등 다양한 활성을 가진 이차 대사산물을 만드는 것이다(Clardy 등, 2006; Hwang 등, 2014). 이러한 특징으로, Streptomyces phaeochromogenes로부터 초파리와 담배거세미나방(Spodoptera littoralis)에 대한 살충능력이 보고된 바 있다(Bream 등, 2001). 또한 셀룰로오즈를 분해하는 특징을 이용하여 해충에 대해 살충능력을 보이는 것으로 보고되었다(Book 등, 2014). 방선균은 직접적인 살충능력뿐 아니라, 파밤나방, 솔나방(Dendrolimus punctatus), 배추좀나방, 콩진딧물(Aphids glycines)과 지하집모기(Culex pipiens) 등의 생장을 방해하는 것으로도 알려져 있다(Table 1; Liu 등, 2008).
방선균으로부터 생성된 살충 물질은 flavensomycin, antimycin A, piericidins, macrotetralides, prasinons 등이 알려져 있으며 S. avermitills로부터 생성된 에버멕틴(avermectins)에 대한 연구가 활발히 진행되어 왔다(Turner와 Schaeffer, 1989). 에버멕틴은 마크로사이클릭 락톤(macrocyclic lactone) 유도체로 알려져 있는데 곤충의 신경시스템 주변에 존재하는 감마-아미노뷰티르산(gamma-aminobutyric acid, GABA)의 수용체를 표적으로 하여 곤충의 신경전달을 방해하고 운동신경계를 마비시킨다(Fig. 2A; Pitterna 등, 2009). 에버멕틴과 유사한 독소로는 나비목의 곤충에게 살충 활성을 나타내는 것으로 알려진 에마멕틴(emamectin)과 S. hygroscopicus Jensen에서 분리된 밀버멕틴(milbemectin) 등이 있다(Ruiu, 2015). Streptomycetes의 또 다른 살충 활성 물질로는 S. aureus, S. albidum 및 S. griseus로부터 각각 분리된 tetranectin, faerifungin, macrotetrolides 등이 보고되어 있다.
한계점: 작물에 대한 방제 방법으로 합성 화학 물질보다는 잔류성이 낮은 생물학적 방제법이 제시되고 있다. 하지만 해충방제를 위한 병원균의 사용은 해충뿐만 아니라 경우에 따라서는 식물에도 유독한 물질(phytotoxin)을 분비하는 경우도 있다. 예를 들어, 방선균이 분비하는 rhizobitoxin (2-amino-4-(2-amino-3-hydroxypropoxy)-trans-but-3-enoic-acid)은 식물의 cystathione-β-lyase 활성을 방해하여 식물체 잎의 황백화(chlorosis)를 일으킨다(Okazaki 등, 2004). 또한 rhizoxin의 경우 β-tubulin에 결합하는 macrocyclic polyketide로, 어린 벼의 세포분열에 영향을 끼치고 식물체를 사멸시킨다(Koga-Ban 등, 1995). 이런 문제점을 해결하기 위하여 생물학적 방제를 위한 미생물의 사용은 표적으로 하는 식물체에 따라서 다르게 적용해야 하며, 병원균 자체를 처리하는 방법 이외에 병원균이 생성하는 살충 물질을 추출하여 사용하는 것도 고려되어야 한다. 다른 문제점은 농업 환경에서 미생물의 생존기간이다. 실제로 포장에서 태양열을 포함하여 온도, 습도, pH 등은 미생물이 선호하지 않는 조건일 수 있기 때문에 작물보호를 위해 작용될 수 있는 미생물의 지속성이 기대 이하로 짧을 수 있다. 이를 극복하기 위하여 사용되는 Bacilli의 경우 불리한 환경에서 포자를 형성하여 살아남을 수 있는 장점을 가지고 있기 때문에 지금까지 많은 제품이 등록될 수 있었다. 마지막으로 Bacilli의 경우에도 보듯이 미생물의 의한 해충방제는 화학 농약과 달리 살충 효과가 나타나기까지 일정시간이 걸리는 것이 문제시될 수 있다. 따라서 미생물의 농도 선정 및 식물체로의 전달이 잘 될 수 있는 방법이 고안된다면 생물학적 방제제로서 이용가치가 높아질 것이다.
진균에 의한 해충의 생물적 방제 방법
해충은 기주로부터 양분을 취하는 방식에 따라 크게 섭식성 해충과 흡즙성 해충으로 구분된다(Fang 등, 2012). 세균이나 바이러스를 이용한 생물학적 방제법은 대부분 기주가 곤충병원성 세균 또는 바이러스를 섭취하여 체내에서 살충 효과를 보이는 것으로 나타나기 때문에 이들을 이용한 방제법은 주로 섭식성 해충에 국한되었다(Fang 등, 2012). 하지만 곤충병원성 진균의 경우 해충 외피의 구성 성분인 큐티클에 직접 침투할 수 있기 때문에 기주와의 접촉만으로도 감염시킬 수 있다(Fang 등, 2012; Gonzalez 등, 2016). 따라서 이러한 곤충병원성 진균을 이용한 생물학적 방제법은 섭식성 해충뿐만 아니라 흡즙성 해충에도 적용이 가능하다(Fang 등, 2012; Gonzalez 등, 2016; Ramanujam 등, 2014).
곤충병원성 진균은 일반적으로 공통된 침입 메커니즘을 갖는 것으로 알려져 있다(Ramanujam 등, 2014). 곤충병원성 진균은 기주 곤충의 표면에 정착한 후 키틴분해효소, 단백질분해효소, 지질분해효소와 같이 다양한 효소를 생성하여 곤충의 큐티클을 분해하고 침입한다(Ramanujam 등, 2014). 침입한 진균은 곤충의 체내에 정착한 후, 기주의 조직으로부터 양분으로 섭취하며 증식함으로써 기주의 내부 조직을 파괴하여 기주를 사멸시킨다(Inci 등, 2014; Malekan 등, 2015). 그 후 균사가 곤충 외피의 얇은 부분을 뚫고 나와 포자를 외부에 비산한다. 대표적인 곤충병원성 진균은 동충하초로, 곤충에 감염되면 외부의 단단한 층과 균사만 남기고 속이 비게 된다(Inci 등, 2014; Malekan 등, 2015).
2014년을 기준으로 상업화된 제품 형태로 사용되는 진균은 170여 개의 제품이 개발되어 있으며, Beauveria bassiana가 전체의 33.9%, Metarhizium anisopliae가 33.9%, Isaria fumosorosea가 5.8%의 비율을 차지하고 있는데 이들은 대부분 동충하초목에 속한다(Ramanujam 등, 2014). 본 단락에서는 곤충 병원성 진균 중 활발히 연구되고 있는 동충하초목 진균의 살충 활성과 병원성 인자에 대하여 소개하고자 한다(Table 1).
① Beauveria bassiana
B. bassiana는 동충하초목 맥각균과 백강균속의 진균으로 1835년 Bassi에 의해 B. bassiana가 누에나방에 백강병(white muscardine)을 일으킨다는 것이 보고되었으며(St. Leger 등, 2011) 가장 대표적으로 사용되는 생물학적 살충제 중 하나이다(St. Leger 등, 2011; Vega 등, 2008). B. bassiana는 기주 범위가 넓어 700종 이상의 곤충을 기주로 가지는 것으로 보고되었다(Xu 등, 2009). 또한 B. bassiana는 유충과 성충의 구분없이 곤충의 여러 성장단계에 감염할 수 있는 특징 때문에(Xu 등, 2009), 담배가루이, 진딧물, 총채벌레(Thrips palmi), 감귤나무이(Diaphorina citri), 바구미(Sitophilus oryzae), 가루깍지벌레(Pseudococcus comstocki) 등에 높은 살충 활성을 나타내는 것으로 보고되었다(Gonzalez 등, 2016). B. bassiana를 가루이에 109 conidia/ml의 농도로 처리했을 때 최대 치사율이 85%로 나타났으며(Malekan 등, 2015; Quesada-Moraga 등, 2006), 배추좀나방 2령 유충에 107 conidia/ml를 처리 시 80%-100%의 높은 치사율을 나타내었다(Duarte 등, 2016; Safavi, 2013).
이러한 높은 살충 효과를 보이는 B. bassiana의 병원성 인자는 저분자의 독성물질인 beauvericin이 처음으로 알려졌고, 이후에 bassianin, bassianolide, beauverolides, beauveriolides, tenellin, oosporein, 옥살산(oxalic acid), bassiacridin 등이 살충 활성을 나타내는 것으로 보고되었다(Fig. 2A; Safavi, 2013). 이러한 저분자 화학 물질 이외에도 꿀벌부채명나방을 모델 시스템으로 사용하여 B. bassiana의 살충 독소 단백질인 Bb70p가 보고되었다(Khan 등, 2016).
위와 같이 B. bassiana는 곤충 병원성 진균 중 가장 넓은 기주 범위를 보이며 높은 살충 효과와 다양한 병원성 인자에 대한 연구가 활발히 수행되고 있기 때문에 곤충 병원성 진균 중 이용 가능성이 매우 높을 것으로 생각된다.
② Metarhizium anisopliae
M. anisopliae는 녹강균속에 속하는 불완전균류로 기존에는 Entomophthora anisopliae라고 알려져 있었으나 최근에는 Metarhizium속으로 재분류되었다(Weeks 등, 2017). 1880년대에 밀풍뎅이(Anisoplia austriac)와 사탕무바구미(Cleonus punctiventris)에 대한 살충 효과가 처음으로 보고되었다(Roberts와 St. Leger, 2004). 그 후, 104 spores/g 정도의 낮은 농도에서도 딸기뿌리바구미(Otiorhynchus ovatus)에 대한 M. anisopliae의 병원성(virulence)이 높게 나타났다(Fisher와 Bruck, 2008). M. anisopliae의 병원성 인자는 destruxins A, B, C, D, E, F가 독소로서 기주 곤충의 체내에서 높은 살충 활성을 나타내는 것으로 보고되어 있다(Fig. 2A; Kershaw 등, 1999).
M. anisopliae는 오랜 기간 연구되어 온 미생물 살충제이며, M. anisopliae는 B. bassiana와 함께 진균을 이용한 생물 살충제로 개발되어 대표적으로 사용되고 있다(Weeks 등, 2017). 또한 살충 효과가 재배지에서도 검증되었기 때문에 실제 농업 현장에서 생물학적 방제로서 적용이 가능할 것으로 생각된다.
③ Lecanicillium muscarium
L. muscarium은 동충하초과에 속하는 곤충 병원성 진균으로 기존에는 Verticillium lecanii로 동정되었으나 2001년 L. muscarium으로 재분류되었다(Gams와 Zare, 2001). L. muscarium의 살충 활성은 주로 성충보다는 알이나 유충에 대하여 더 높은 활성을 나타내는 것으로 보고되었으며(Duarte 등, 2016), bassianolid와 dipicolinic acid가 병원성 인자로 알려져 있다(Jirakkakul 등, 2008; Kulkarni, 2015).
L. muscarium을 유기 합성 살충제와 함께 처리하면 담배가루이 유충에 대하여 81%-89%의 치사율을 나타냈고 L. muscarium을 단독으로 처리했을 시에는 24시간 후 알에서 55%, 유충에서 68%의 치사율을 나타내는 것으로 보고되었다(Cuthbertson 등, 2005, 2008; Duarte 등, 2016). 기내(in vitro) 실험에서 L. muscarium 107 conidia/ml를 차 잎에 처리하고 거세미나방(Rhyacia simulans)의 유충에게 경구 투여했을 때, 7일 후 50%-75%의 치사율을 나타내었다(Güçlü, 2010). 포장 실험에서도 참다래에 L. muscarium Lm4를 107 conidia/ml 처리한 후 거세미나방의 유충과 성충에 경구 투여 시 살충 활성을 나타내는 것으로 보고되었으며, 거세미나방의 경우 유충이 성충에 비해 L. muscarium에 대해 감수성을 나타내는 것으로 보고되었다(Güçlü 등, 2010). 온실가루이(Trialeurodes vaporariorum) 유충의 초기와 후기에 L. muscarium을 103-106 conidia/ml로 분무하였을 때 각각 62%, 87%의 치사율을 나타내어 온실가루이의 경우 초기의 유충에게 처리하는 것이 방제 효과가 증대되는 것으로 보고되었다(Malekan 등, 2015).
L. muscarium은 기내 실험과 포장 실험에서의 적용 효과에 관한 연구가 보고되었으나, 곤충 성장단계에 따라 L. muscarium에 대한 감수성 차이로 인해 실제 현장에 적용하기 위해서는 진균 살충제를 처리하는 시기에 관한 연구가 필요하다고 생각된다.
④ Isaria fumosorosea
I. fumosorosea는 1904년 우크라이나의 바구미에서 분리된 곤충병원성 진균으로 Trichocomaceae의 Paecilomyces fumosoroseus로 알려졌으나, 2012년 자낭의 형성과정 및 형태에 따라 Cordycipitaceae에 속하는 I. fumosorosea로 재분류되었다. I. fumosorosea는 최근 생물학적 방제제로 알려지기 시작하여 가루이와 같은 해충의 방제에 사용되고 있다(Zimmermann, 2008). I. fumosorosea는 감귤녹화병(citrus greening disease)을 일으키는 감귤나무이와 감귤진딧물(Toxoptera citricida), 유리날개나방(Homalodisca vitripennis) 같은 감귤류 해충에 살충 활성을 나타내는 것으로 보고되었다(Avery 등, 2016; Casique-Valdés 등, 2015; Hunter 등, 2011). 감귤뿌리바구미(Diaprepes abbreviatus)의 경우 I. fumosorosea를 분무 처리하고 7일 후 유충과 성충에서 각각 13%, 19%의 치사율을 나타내었으며, 감귤뿌리바구미 성충에게 I. fumosorosea 포자를 처리한 잎을 경구 투여 시 35일 후 100% 치사시키는 것으로 보고되었다(Avery 등, 2016).
I. fumosorosea의 살충 메커니즘과 병원성 인자에 대해서는 아직 연구가 부족하지만, 최근 키틴분해효소 유전자 Ifchit1이 I. fumosorosea의 병원성에 중요한 역할을 한 것으로 알려졌다(Huang 등, 2016). I. fumosorosea를 이용한 생물학적 방제법은 감귤류 작물에서 이미 포장 적용 실험을 통한 실증효과가 검증되어 현장에서 이용하고 있다. 그러나 다양한 작물에 적용하기 위해서는 살충 메커니즘이나 병원성 인자에 대한 특성연구가 필요할 것이라 생각된다.
한계점: 곤충병원성 진균의 경우 대량의 포자를 형성하기 때문에 세균이나 바이러스보다 제제화와 대량 생산이 용이하고, 작용 메커니즘이 화학 농약에 비해 다양하여 곤충이 저항성을 갖기 어렵다는 장점이 있다(Haase 등, 2015; Kergunteuil 등, 2016). 하지만 진균을 이용한 방제법은 다른 생물 농약에 비해 살충 활성이 나타나기까지의 시간이 길고, 처리 시 접종농도가 높아야 하며 환경 스트레스에 취약하며 인해 낮은 살충 활성을 나타내 제품개발을 위한 연구가 부족한 실정이다(Fang 등, 2012; Lacey 등, 2015). 이러한 단점을 극복하기 위해 멜라닌 유전자를 삽입하는 등의 다양한 연구가 이루어지고 있으며(Fang 등, 2012; St. Leger 등, 2011; Tseng 등, 2011) 처리 방식의 최적화 연구를 통해 단점들을 보완한다면 생물학적 살충제로서 이용가치가 높을 것으로 예상된다.
바이러스 의한 해충의 생물적 방제 방법
곤충병원성 바이러스는 1960년대부터 보고되어 왔으며 현재까지 650여 개의 곤충병원성 바이러스가 곤충으로부터 분리되었다(Moscardi 등, 2011; Popham 등, 2016). 주로 나비목과 벌목의 유충에 대하여 살충 효과를 나타내며(Inci 등, 2014), 높은 기주 특이성을 나타내는 것으로 보고되었다(Ramanujam 등, 2014).
곤충병원성 바이러스에 의한 살충 메커니즘은 기주에 침입한 바이러스가 세포 내에서 복제 과정에 의해 일어나는데, 일반적으로 기주의 섭식에 의해 장내로 침입하여 감염되고(Shim 등, 2013), 곤충의 교배기나 산란기 동안 곤충 간에 전염이 일어나기도 한다(Inci 등, 2014). 바이러스에 감염된 기주는 움직임이 둔해지고 표면의 큐티클이 창백하게 변색되는 병징을 보이는데, 병징이 나타난 후 1-2일 사이에 기주의 세포가 분해되어 바이러스가 외부로 방출된다(Inci 등, 2014). 한편, 곤충병원성 세균이나 진균과 다르게 곤충병원성 바이러스의 경우, 자체적으로 생산하는 독소에 의한 살충 효과는 크지 않았으나 외부의 독소 생성 유전자를 삽입하여 살충 효과를 높이려는 연구가 90년대부터 현재까지 이루어지고 있다(Haase 등, 2015; Shim 등, 2013).
곤충병원성 바이러스는 대부분 dsDNA (double-stranded DNA) 바이러스 속의 Baculoviridae에 속하는 것으로 알려져 있기 때문에(Kergunteuil 등, 2016; Popham 등, 2016) 본 단락에서는 대표적인 곤충병원성 바이러스인 배큘로바이러스(baculovirus)를 중심으로 이야기하고자 한다.
① Baculovirus
1947년 누에나방의 배큘로바이러스 감염이 최초로 보고된 이후, 알팔파자벌레(Autographa californica)에 감염하는 Autographa californica NPV를 포함한 100여 개 종의 배큘로바이러스가 보고되었다(Table 1; Bonning과 Nusawardani, 2007). 배큘로바이러스에 의한 살충 활성은 주로 담배나방(Helicoverpa)속이나 밤나방(Spodoptera)속과 같은 나비목 해충에서 연구되었는데(Lacey 등, 2015; Rowley 등, 2011), 합성 살충제에 대하여 빠르게 저항성을 획득하는 나비목의 해충을 방제하기 위하여 배큘로바이러스를 포장에 적용하기 위한 연구가 주목받아 왔다(Arrizubieta 등, 2014; Lacey 등, 2015).
배큘로바이러스는 바이러스 중에서는 비교적 크기가 크고 복잡한 구조를 가지고 있다. 가장 외부에는 과립형(granule) 혹은 다면체(polyhedral) 형태의 단백질로 구성된 폐색체(occlusion body)를 가지고 있다(Fig. 3A). 배큘로바이러스는 폐색체의 형태에 따라 NPV (Nucleopolyhedrovirus)와 GV (Granulovirus)로 분류된다(Fig. 3A; Lacey 등, 2015). 폐색체 내부에는 막대모양으로 생긴 핵단백질체(nucleocapsid)가 80-180 kb 정도 크기의 dsDNA 유전체를 감싸고 있으며(Ramanujam 등, 2014), 폐색체 내부의 구조를 통틀어 비리온(virion)이라고 한다(Lacey 등, 2015). 이와 같은 배큘로바이러스의 구조적인 특징 때문에 유전물질이 노출되어 있는 다른 바이러스보다 유전물질을 보호하기 용이하고, 제품화하는 경우에 조제와 적용방법, 장기 보관에 유리할 것으로 생각된다.
Fig. 3
Classification of Nucleopolyhedrovirus (NPV) and Granulovirus (GV) and infection cycle of Baculoviruses. (A) Baculoviruses has two genera which are characterized by morphology of occlusion bodies. NPV has multiple virions in polyhedral form occlusion bodies while GV has single virion in granule form. NPV strains defined as having a single or multiple nucleocapsid in an envelope. (B) Infection cycle of baculoviruses. 1. Baculoviruses are ingested by insect larvae. 2. The occlusion bodies are dissolved in the alkaline mid-gut and virions infects the host through the mid-gut cells. Virus particles then replicate inside the insect cells. 3. Virus particles are released to the environment when infected cells were disintegrated.

배큘로바이러스가 기주에 침투하면 장 내부에 도달한 후 바이러스의 DNA를 감싸고 있는 비리온이 해체되면서 유충의 중장 세포를 감염하게 된다(Fig. 3B; Kergunteuil 등, 2016; Moscardi 등, 2011). 감염된 기주의 세포 내에서 기주의 세포활동을 저해하는 키틴분해효소 등을 생성하여 내부 구조를 파괴하고 기주를 사멸시킨다(Kroemer 등, 2015).
NPV (Nucleopolyhedrovirus): 브라질에서는 대두에 감염하는 Anticarsia gemmatalis를 방제하기 위하여 AngeNPV (Anticarsia gemmatalis NPV)를 사용하고 있으며(Moscardi, 1999), 중국에서는 면화다래벌레(Helicoverpa zea)를 방제하기 위하여 HearNPV (Helicoverpa armigera NPV)를 실제 재배지에서 이용하고 있다(Bonning과 Nusawardani, 2007; Sun, 2015). 이 외에도 Mamestra brassicae NPV는 도둑나방(Mamestra brassicae), 파밤나방, 담배거세미나방 방제에 주로 사용하는 것으로 보고되었다(Sun, 2015).
NPV를 바이러스 살충제로서 이용하기 위한 연구가 오랫동안 이루어져 왔으며, 살충제로서 배큘로바이러스의 단점이라고 알려진 살충 효과가 나타나기까지의 시간이 화학농약에 비해 길다는 점을 보완하고자 전갈이나 진드기, 거미 등의 독소 생성 유전자를 바이러스에 삽입하는 연구들이 이루어졌다(Haase 등, 2015; Kergunteuil 등, 2016; Shim 등, 2013). AcMNPV (Autographa californica NPV)를 이용한 유전자 재조합 연구에서 궐련벌레살이주머니응애(Pyemotes tritici)의 독소 유전자인 tox34를 삽입한 경우 양배추은무늬밤나방(Trichoplusia ni)과 가을조밤나방(Spodoptera frugiperda) 유충에 대해서 살충 활성을 나타내는 시간이 기존 바이러스에 비해 50%-60%로 감소하여 야생형 바이러스에 비해 빠른 살충 활성을 나타내는 것으로 보고되었다(Regev 등, 2003). 최근에는 독거미(Lachesana tarabaevi)의 cyto-insectotoxin 유전자를 이용한 연구도 진행되고 있다(Ali 등, 2015; Kergunteuil 등, 2016). 그 밖에도 높은 살충 활성을 나타내는 것으로 알려진 곤충병원성 세균 B. thuringiensis의 Cry 독소 생성 유전자를 곤충병원성 바이러스 AcMNPV에 삽입한 후 파밤나방 유충에 처리 시 치사시간이 약 50% 감소한 것으로 알려졌다(Shim 등, 2013). 상기의 연구들을 통하여 NPV의 단점인 살충 효과의 발현시간이 보완된다면 효율적인 생물학적 방제제로 이용이 가능해질 것으로 생각된다.
GV (Granulovirus): Cydia pomonella GV는 사과 작물의 대표적인 해충인 코들링나방(Cydia pomonella)의 방제제로, 유럽에서 1987년 개발되어 최근에도 사용되는 바이러스 살충제이다(Lacey 등, 2015). 이외에도 배추좀나방, 감귤코들링나방(Cryptophlebia leucotreta), 애모무늬잎말이나방(Adoxophyes orana), 배추흰나비(Pieris rapae) 방제에도 살충 효과를 나타내는 것으로 알려져 있다(Lacey 등, 2015). 최근에는 PoGV (Phthorimaea operculella GV)를 식물에 처리 시 지하부의 해충에 대해 살충 활성을 나타낸다는 것이 보고되었으며(Kergunteuil 등, 2016), 실제로 토양 처리 시 감자뿔나방(Phthorimaea operculella) 유충의 개체수가 73% 감소하는 것으로 보고되었다(Kergunteuil 등, 2016).
② 기타 살충 활성을 나타내는 바이러스
폐색체가 없는 Nudiviridae의 한 종인 Oryctes virus는 유야자수(oil palm) 해충인 남방장수풍뎅이(Oryctes rhinoceros)에 대해 살충 활성을 나타내었으며(Lacey 등, 2015; Ramle 등, 2005), 외피단백질이 이중, 삼중으로 되어있는 Reoviridae에 속하는 Cypovirus는 유야자수의 해충 방제에 대한 효과가 보고되었으나 상용화되지는 않았다(Lacey 등, 2015; Zeddam 등, 2003). 현재까지 배큘로바이러스 이외의 바이러스가 나타내는 살충 활성에 대한 연구는 활발하지 않으며 심각한 병해를 입히는 해충에 대하여 특별한 살충 효과를 보이지 않는 한 배큘로바이러스를 이용한 생물학적 방제제를 개발하는 것은 어려울 것으로 보인다(Lacey 등, 2015).
한계점: 곤충병원성 바이러스의 경우, 생물학적 방제제로서 대량 생산이 어렵기 때문에 상업화가 세균과 곰팡이에 비해 상대적으로 어렵다. 특히 NPV는 살아있는 기주에서만 증식이 가능하기 때문에 인공 미생물 배지에서는 배양할 수 없다. 따라서 NPV를 생물학적 방제제로서 개발하기 위해서는 적합한 곤충 세포주(cell line)에 대한 연구가 선행되어야 한다(Ramanujam 등, 2014). 현재까지 배큘로바이러스를 배양하기에 적합한 세포주가 다수 개발되었으나 바이러스는 세균이나 효모 세포에 비해 분해되기 쉬워 실제 대량 생산까지는 개발되지 못했다(Lacey 등, 2015). 따라서 생물학적 방제제로서 곤충병원성 바이러스의 안정성을 높이기 위해서는 10톤 이상의 대량 생산이 가능한 곤충 세포주를 개발하는 것이 중요하다.
선충의 생물적 방제 방법
선충 방제 미생물은 선충포식성 진균과 선충병원성 세균이 있는데 이들 미생물은 선충 표면에 부착하여 표면을 분해하거나, 장에서 독소물질을 생성한다(Li 등, 2015). 선충병원성 미생물들은 키틴분해효소, 단백질분해효소, 콜라겐분해효소를 분비하여 선충의 표피와 알 표면에 침투하여 외벽을 파괴하고, 대사물질이나 단백질을 외부로 분비하여 선충의 생리와 생장을 억제한다(Li 등, 2015). 최근 이러한 선충병원성 미생물을 이용한 친환경적인 선충 방제제의 개발이 증가하고 있다. 본 단락에서는 살선충 활성을 나타내는 진균과 세균을 소개하고, 이들이 선충의 치사를 이끌어내는 메커니즘에 대해 설명하고자 한다.
세균에 의한 선충의 생물학적 방제 방법
선충병원성 세균은 토양 내에 존재하여 자연계에서 동식물 기생성 선충의 포식자 역할을 하고 있다. 선충병원성 세균은 선충과의 상호작용 측면에서 절대기생균, 기회기생균, 근권세균, Cry 독소 생성균, 내생균, 공생균으로 분류할 수 있다(Tian 등, 2007). 본 단락에서는 선충병원성 세균 중 대부분을 차지하는 Bacillus, Pseudomonas, Pasteuria속 세균과 이외의 살선충 활성을 나타내는 세균에 대하여 소개하고자 한다(Table 2).
Table 2
Microbes, their microbial determinants, and the commercial products for controlling the plant pathogenic nematodes
| Species | Target insects | Effector/determinant | Products | References |
|---|---|---|---|---|
| Bacteria | ||||
| Pasteuria penetrans | Meloidogyne spp. | Endospore; Velcro-like mechanism of attachment | Econem | Noel et al,. 2005; Persidis et al., 1991 |
| P. thornei | Pratylenchus spp. | |||
| P. nishzawae | Heterodera spp.; Globodera spp. | |||
| Pseudomonas fluorescens | Xiphinema americanum; G. rostochiensis; Meloidogyne spp.; Helicotylenchus multicinctus | 2,4-Diacetylphloroglucinol; Protease AprA; Hydrogen cyanide | Sudozome, Nemato-kill | Cronin et al., 1997; Meyer et al., 2009, 2016; Neidig et al., 2011; Selvaraj et al., 2014; Siddiqui and Shaukat, 2003a, 2003b, 2004a; Siddiqui et al., 2005, 2006 |
| P. aeruginosa | Caenorhabditis elegans | Phenazine-1-carboxylic acid; 1-Hydroxyphenazine; Pyocyanin; Pyoverdin | - | Cezairliyan et al., 2013; Kirienko et al., 2013; Mahajan-Miklos et al., 1999; Slininger and Shea-Wilbur, 1995; Van Rij et al., 2004; |
| Bacillus thuringiensis | Meloidogyne spp.; Pratylenchus spp.; Heterodera spp. | Cry toxin; Metalloproteinase Bmp1 | Tobagi, BT1, Chungsanyang, Geumulmang, Solbichchae, Bigyeol, Eco-zone | Schnepf et al., 1998 |
| B. nematocida | Panagrellus redivivus; Bursaphelenchus xylophilus | Benzaldehyde; 2-Heptanone; Benzyl benzoate; Acetophenone; Alkaline serine protease Bace16; Neutral protease Bae16 | Nemix, Biostart, BioYield | Deng et al., 2013; Huang et al., 2005b; Niu et al., 2010 |
| B. megaterium | M. exigua | Benzeneacetaldehyde; 2-nonanone; Decanal; 2-Undecanone; 2,3-Dimethyl-butanedinitrile; 3,5-Dimethoxytoluene; Dimethyl disulphide | Huang et al., 2010; Oliveira et al., 2007 | |
| B. firmus | Meloidogyne spp.; H. glycines; Ditylenchus dipsaci; Radopholus similis | Serine protease Sep1 | Lian et al., 2007; Zheng et al., 2016a, 2016b | |
| B. cereus | Meloidogyne spp. | Sphingomyelinase C; Cry4Aa; Sphinganine; Phytosphingsine | Gao et al., 2016; Nishiwaki et al., 2004; Poopathi et al., 2014 | |
| B. pumilus | Meloidogyne spp. | Chitinase; Serine protease | Akhtar and Siddiqui, 2008; Lee and Kim, 2016; Oliveira et al., 2007; Ramezani et al., 2014 | |
| Brevibacillus laterosporus | H. glycines; B. xylophilus | Alkaline serine protease BLG4 | - | Decraemer et al., 2003; de Oliveira et al., 2004; Huang et al., 2005a; Tian et al., 2006 |
| Fictibacillus phosphorivorans | C. elegans; Meloidogyne spp. | Amidophos-phoribosyl transferase PurL; Alkaline serine protease BLG4; Calcium-transporting ATPase Eca1 | - | Fan et al., 2007; Huang et al., 2005a; Xia et al., 2011; |
| Streptomyces avermitilis | M. incognita | Avermectin | AVICTA®, Doctor-farm, Sol-vaccine, Index | Cabrera et al., 2013 |
| Streptomyces sp. CMU-M021 | Fervenuline; Isocoumarin | Ruanpanun et al., 2011 | ||
| S. albogriseolus | Fungichromin B | Zeng et al., 2013 | ||
| S. hydrogenans DH16 | 10-(2,2-dimethyl-cyclohexyl)-6,9-dihydroxy-4,9-dimethyl-dec-2-enoic acid methyl este (SH2) | Kaur et al., 2016 | ||
| Serratia mercescens | Meloidogyne spp.; Tylenchulus semipenetrans | Prodigiosin | - | Suryawanshi et al., 2014 |
| Alcalinenes faecalis | Meloidogyne spp. | Serine protease Esp | - | Ju et al., 2016 |
| Fungi | ||||
| Arthrobotrys oligospora, A. dactyloides, A. conoides | Meloidogyne spp.; Pratylenchus spp.; Heterodera spp. | Serine protease PII, AozI | - | Li et al., 2015; Meerupati et al., 2013; Tucker and Talbot, 2001 |
| Drechmeria coniospora | Meloidogyne spp. | Proteases; Heat-labile enterotoxin | - | Lebrigand et al., 2016; Moosavi and Zare, 2012 |
| Esteya vermicola | Bursaphelenchus spp.; Aphelenchoides besseyi; A. fragariae; A. ritzemabosi; D. angustus; D. dipsaci; D. destructor; Rhadinaphelenchus cocophilus; M. incognita; H. avenae; P. penetrans | α-Pinene; β-Pinene; Camphor; Chitinase | - | Kubátová et al., 2000; Liou et al., 1999; Wang et al., 2008, 2014 |
| Pochonia rubescens, P. chlamydosporia | G. pallida | Serine protease P32, VCP1; Chitinase Chi43 | Xianchongbike | Lopez-Llorca, 1990; Segers et al., 1994; Tikhonov et al., 2002; Yang et al., 2007a |
| Paecilomyces lilacinus | M. incognita; G. rostochiensis | Leucinotoxins; Serine protease pSP-3; Chitinase Plc | Bioact, NemOut, PROPHYTA | Brand et al., 2004; Khan et al., 2006; Kiewnick and Sikora, 2006; Park et al., 2004 |
| Talaromyces thermophilus | M. incognita; B. siylopilus | Thermolides | - | Guo et al., 2012 |
| Coprinus comatus | M. incognita | 5-Methylfuran-3-carboxylic acid; 5-Hydroxy-3,5-dimethylfuran-2(5H)-one | - | Luo et al., 2007 |
| Tricoderma spp. | Meloidogyne spp. | Trypsin like protease PRA1; Serine protease SprT; Chitinase chi18-5, chi18-12; Trichodermin; β-Vinylcyclopentane-1α,3α-diol; 6-Pentyl-2H-pyran-2-one; 4-(2-Hydroxyethyl) phenol | Jayeon-ilang, Neff Kill | Suarez et al., 2004; Szabó et al., 2012; Vachon et al., 2012; Yang et al., 2010 |
| Myrothecium verrucaria | Meloidogyne spp.; Pratylenchus spp.; Heterodera spp. | Protase; Chitinase | DiTera® | Dong et al., 2015; Wilson and Jackson, 2013 |
| Aspergillus niger F22 | M. hapla | Oxalic acid | Nema-free | Jang et al., 2016 |
① Pasteuria spp
Pasteuria속은 선충의 절대기생균으로 균사와 내생포자를 형성하고, 식물기생성 선충과 갑각류에 기생하는 그람-양성 세균이다(Tian 등, 2007). Pasteuria spp.를 식물기생성 선충을 방제하기 위한 생물학적 방제제로 이용할 수 있는 이유는 식물기생성 선충을 포함한 300여 종의 선충에 감염할 수 있고 살선충 효과가 안정적이기 때문이다(Bird 등, 2003; Chen과 Dickson, 1998). Pasteuria penetrans는 Meloidogyne spp.와 같은 뿌리혹선충에 기생하고, P. thornei는 Pratylenchus spp.와 같은 뿌리썩이선충에, P. nishzawae는 Heterodera spp., Globodera spp.와 같은 시스트선충에 기생하는 것으로 알려져 있다(Noel 등, 2005).
Pasteuria spp.의 포자는 선충의 2령 유충 표면의 큐티클에 점착하고 발아관(germ tube)을 이용해 선충 내부로 침투한다. 선충의 내부에서 조직을 분해하고 이후 증식한 포자를 토양으로 방출한다(Mankau 등, 1976; Sayre와 Wergin, 1977). Pasteuria spp.는 선충의 큐티클에 점착하기 위해 포자 표면에 β-mercaptoethanol을 포함하는 당단백질로 구성된 섬유소를 생성하는 것으로 알려졌다(Persidis 등, 1991). 이를 생성하지 못하는 Pasteuria spp.의 돌연변이는 선충의 큐티클에 점착하지 못하기 때문에, 이 섬유소가 Pasteuria spp.의 선충 표면 점착에 있어 중요한 인자임을 알 수 있다. Pasteuria spp.의 섬유소는 선충의 큐티클에 존재하는 렉틴 수용체와 결합하여 복합체를 만드는 것으로 보고되었지만 그 메커니즘은 아직 밝혀지지 않았다(Persidis 등, 1991).
② Pseudomonad
P. fluorescens, P. chlororaphis, P. protegens 등은 토양, 해양 환경 등에서 유기물을 분해하거나 식물 근권에 정착하여 생장을 촉진시키는 효과를 나타내지만, P. aeruginosa, P. syringae 등은 각각 사람이나 식물에 감염하는 병원균으로도 알려져 있다(Flury 등, 2016). 1999년에 P. aeruginosa PA14가 예쁜꼬마선충(Caenorhabditis elegans)에 살선충 활성을 나타내는 것이 보고된 이후, Pseudomonas속 세균과 선충 간의 상호작용과 살선충 메커니즘을 규명하기 위한 연구들이 수행되었고, 살선충 활성을 나타내는 독소, 분해효소, 2차대사물질에 대하여 알려졌다(Li 등, 2015). 이 단락에서는 최근 연구를 통해 밝혀진 Pseudomonad의 병원성 인자와 독소, 세포 외 효소에 대해 다루고자 한다(Table 2, Fig. 2B).
HCN (hydrogen cyanide): HCN은 Pseudomonas속 세균이 생산하는 휘발성 화합물로(Blumer와 Haas, 2000), P. protegens CHA0가 생산하는 HCN을 자바니카뿌리혹선충에 처리 시 살선충 활성을 나타내는 것으로 보고되었다. HCN은 미토콘드리아의 사이토크롬산화효소(cytochrome oxidase)를 억제하는 것으로 알려져 있으며(Blumer와 Haas, 2000; Gallagher와 Manoil, 2001), HCN을 생산하는 P. chlororaphis O6을 당근뿌리혹선충에 처리 시 선충의 유충을 치사시키고 알 부화율을 감소시키는 것으로 나타나(Siddiqui 등, 2006) P. chlororaphis O6과 같은 Pseudomonas속 균주가 생산하는 HCN이 주요 살선충 물질로 보고되었다(Lee 등, 2011).
DAPG (2, 4-diacetylphloroglucinol): P. fluorescens가 생산하는 DAPG는 식물병원균의 생장을 저해하며 곤충과 선충의 생육을 억제하는 물질로 작용한다고 알려져 있다(McSpadden Gardener, 2007; Weller, 2007). 게다가 식물 근권 세균으로서 기주 식물에 식물 병원균에 대한 저항성을 유도하고 작물의 생산량을 증대시키는 것으로 보고되었다(Iavicoli 등, 2003; Siddiqui와 Shaukat, 2003b, 2004b; Van Loon과 Bakker, 2006; Weller 등, 2012). P. fluorescens가 생산하는 DAPG는 미국단검선충(Xiphinema americanum)의 성충에 대해 살선충 활성을 나타내며(Meyer 등, 2009), 감자몬스트포낭선충(Globodera rostochiensis) 유충의 운동성을 억제하고 치사시키는 것으로 보고되었다(Cronin 등, 1997; Meyer 등, 2016). 온실 실험에서 P. protegens CHA0의 배양액을 처리 시 자바니카뿌리혹선충(Meloidogyne javanica), 고구마뿌리혹선충(M. incognita)의 알 부화율이 감소하고 유충을 사멸시키는 것으로 보고되었으며(Meyer 등, 2009; Siddiqui와 Shaukat, 2003a, 2003b, 2004a, 2004b), 콩과 작물(가지, 녹두, 콩)에 P. protegens CHA0의 배양액을 처리시 뿌리혹선충의 뿌리혹 형성이 감소하였다(Meyer 등, 2009; Siddiqui와 Shaukat, 2003a, 2003b). 또한 P. fluorescens Pf-1도 바나나 재배지에서 바나나나선선충(Helicotylenchus multicinctus)의 감염을 억제하는 것으로 나타나(Meyer 등, 2016; Selvaraj 등, 2014), Pseudomonas spp.가 생산하는 DAPG를 토양에 처리하면 식물기생성 선충의 감염이 감소하는 것을 알 수 있었다(Meyer 등, 2016; Neidig 등, 2011; Siddiqui 등, 2005, 2006). Pseudomonas spp.의 DAPG를 생성하는 유전자는 phlHGFACBDE의 8개의 유전자로 구성되어 있으며, phlD가 type III polyketide synthase로 DAPG를 생성하는 데 중요한 유전자로 알려졌다(Bangera와 Thomashow, 1999).
AprA (alkaline protease): P. fluorescens의 염기성 단백질분해효소 AprA는 고구마뿌리혹선충의 알 부화를 억제하고 유충을 마비시켜 살선충 활성을 나타내는 것으로 보고되었다(Siddiqui 등, 2005). P. aeruginosa, P. fluorescens 등의 유전체 분석과 유전자 결손 연구를 통하여, Pseudomonas속 세균의 유전체에는 AprA와 유사한 유전자가 존재하는 것으로 확인되었으며 Pseudomonas spp.의 주요 조절인자로 알려진 GacA가 AprA와 같은 분해효소의 발현을 조절하는 것으로 보고되었다(Guo 등, 2016; Siddiqui 등, 2005). P. fluorescens가 생산하는 단백질분해효소 AprA를 생물학적 살선충제로 이용하기 위해서는 AprA의 살선충 특성과 살선충 작용 메커니즘에 대한 연구가 수행되어야 할 것이다.
페나진(phenazines): P. aeruginosa PA14가 생성한 페나진(phenazine, phenazine-1-carboxylic acid, 1-hydroxyphenazine), pyocyanin은 예쁜꼬마선충 장내 세포에 작용하여 24시간 내에 사멸시킨다(Cezairliyan 등, 2013; Mahajan-Miklos 등, 1999). 페나진은 3환식의 방향성 분자로, P. aeruginosa 외에도 P. fluorescens가 phenazine-1-carboxylic acid를, P. chlororaphis가 phenazine-1-carboxamide를 생성하는 것으로 알려져 Pseudomonas속 세균이 페나진과 유사한 독소를 생성하여 살선충 활성을 나타내는 것으로 보고되었다(Slininger와 Shea-Wilbur, 1995; Van Rij 등, 2004). P. aeruginosa의 독소 생산과 관련된 유전자는 phzABCDEFG의 7개 유전자 집단으로 구성되어 있으며(Mavrodi 등, 2001) 페나진 생산을 조절하는 메커니즘과 살선충 메커니즘에 대한 연구가 수행되고 있다.
Pyoverdin: P. aeruginosa PA14는 독소물질인 페나진뿐만 아니라, Pseudomonas spp.가 생성하는 사이드로포어(siderophore)인 pyoverdin도 살선충 활성을 가진다(Kirienko 등, 2013). Pyoverdin은 예쁜꼬마선충에 Pseudomonas spp.가 감염되면 선충의 세포 내 철분 항상성을 무너뜨려 저산소증으로 사멸하게 하는 것으로 보고되었다(Kirienko 등, 2013). 그러나 페나진에 비해 Pseudomonas spp.와 선충 간의 상호작용 및 치사 메커니즘에 대한 연구가 미비한 편이다.
③ Bacilli
1972년 B. thuringiensis를 처리 시 뿌리혹선충의 밀도가 감소한 것으로 보고된 이후, 친환경 생물학적 선충 방제제를 개발하기 위해 B. thuringiensis와 유사한 살선충 활성을 나타내는 미생물에 대한 보고가 이어졌다. Bacillus속은 이전부터 식물병원균을 억제할 뿐만 아니라 식물의 근권에 정착하여 기주 식물의 저항성을 유도하는 것으로 알려져 있다(Ongena와 Jacques, 2008). 또한 뿌리혹선충과 같은 식물기생성 선충과 각종 충해를 방제할 수 있다고 보고되기도 했다. B. cereus (Oka 등, 1993), B. firmus (Mendoza 등, 2008; Terefe 등, 2009), B. subtilis (Xia 등, 2011), B. nematocida (Niu 등, 2007), B. megaterium (Huang 등, 2010)과 같은 다양한 Bacillus속 균주들이 살선충 활성을 나타내는 것으로 밝혀져 있어 각 균주에 대한 특성과 살선충 효과에 대해 소개하고자 한다(Table 2, Fig. 2B).
B. thuringiensis: B. thuringiensis는 포자형성 중에 Cry 독소라는 내독소를 생성한다(Schnepf 등, 1998). Cry 독소는 애벌레, 초시류 등의 각종 곤충과 선충에 독성을 나타내고 척추동물에는 영향을 끼치지 않는 이상적인 생물학적 방제제로 보고되었다. Cry 독소가 생물학적 방제제로 주목 받기 시작한 이후, 다양한 Cry 독소 생성 유전자를 식물에 삽입하여 선충 피해에 대한 방제법으로 사용하려는 시도가 있었다(Li 등, 2007b; Sanahuja 등, 2011). 또한 Cry 5B 등 다양한 Cry 독소가 보고되었지만(Griffitts 등, 2005; Vachon 등, 2012), 작물의 적용 후 태양열이나 식물에서 나오는 다양한 화학 성분 등에 의해서 효과가 길게 지속되지 못하기 때문에 농업적 사용이 제한되었다(Raymond 등, 2010). B. thuringiensis에 대해서는 이미 많은 부분이 알려졌기 때문에 본 단락에서는 이외의 다른 선충병원성 세균에 대해 논하고자 한다.
B. nematocida: 내생포자를 생성하는 그람-양성 세균으로 중국의 삼림토양에서 최초로 분리되었고 Panagrellus redivivus와 소나무재선충(Bursaphelenchus xylophilus)에 살선충 활성을 나타내었다(Huang 등, 2005b). B. nematocida는 “트로이 목마 메커니즘”을 이용하여 선충에 감염한다(Niu 등, 2010). B. nematocid의 트로이 목마 메커니즘이란, B. nematocid가 선충이 선호하는 휘발성 물질을 분비하여 선충을 유인하고 선충 내부로 침투한 후 단백질 분해효소를 분비하여 표면과 장을 분해하는 과정을 말한다. B. nematocida가 선충을 유인하기 위해 분비하는 휘발성 화학물질은 benzaldehyde, 2-heptanone, benzyl benzoate, acetophenone으로 선충이 B. nematocida를 먹이로 착각하게 한다(Niu 등, 2010). 선충이 B. nematocida를 섭취하면 B. nematocida는 선충 내부에서 기생하면서 염기성 세린 단백질 분해효소 Bace16과 중성 단백질 분해효소 Bae16을 생성하여 내부조직을 분해하여 선충을 치사시킨다. 2013년에는 B. nematocida의 병원성이 Bacillus spp.의 정족수 인식(quorum sensing) 시스템에 의해 조절되는 것으로 보고되었는데, B. nematocida의 정족수인식 시스템의 ComP 유전자를 결손시킨 돌연변이 균주는 선충을 유인하지 못하고 병원성 인자의 발현이 감소하여 단백질분해효소 Bace16과 Bae16을 생성하지 못하는 것으로 확인되었다(Deng 등, 2013).
B. megaterium: B. megaterium은 1884년 처음 동정된 이후, 내생 포자를 생성하는 그람-양성 세균으로 식물 근권과 소 분변, 곤충 장 등 다양한 환경에서 분리되고 있다. B. megaterium은 식물 근권에 정착하고 기주 식물의 인 가용능을 향상시켜 뿌리의 생장을 촉진하는 것으로 알려져 있어 생물학적 비료로 사용되기도 했다(Çakmakçi 등, 2006; de Freitas 등, 1997). 또한 B. megaterium은 커피뿌리혹선충(Meloidogyne exigua)의 뿌리혹 형성과 알 부화를 감소시키는 것으로 보고되었다(Oliveira 등, 2007). 최근 연구에서는 B. megaterium이 생성하는 살선충 활성을 지닌 휘발성 물질(benzeneacetaldehyde, 2-nonanone, decanal, 2-undecanone, dimethyl disulphide, phenyl ethanone, nonane, phenol, 3,5-dimethoxytoluene, 2,3-dimethyl-butanedinitrile and 1-ethenyl-4-methoxy-benzene)이 보고되었는데, 모두 55%-94%의 살선충 활성을 나타내는 것으로 확인되었다(Huang 등, 2010). B. megaterium은 온실 실험에서도 살선충 활성을 지속적으로 나타내면서 작물의 수확률을 향상시키는 것으로 나타났지만 산업적 적용을 위해서는 대량 배양과 제제화 연구가 필요하다.
B. firmus: B. firmus는 그람-양성의 내생포자를 생성하는 세균으로서 1933년 동정된 이후 다양한 환경에서 분리되고 있으며(Terefe 등, 2009), 중금속 분해 등 다양한 분해효소가 산업적으로 유용하게 적용되고 있다. B. firmus는 뿌리혹선충, 대두시스트선충(Heterodera glycines), 마늘구근선충(Ditylenchus dipsaci), 감귤선충(Radopholus similis)의 유충을 마비시키고 알 부화를 억제하는 것으로 보고되었다(Lian 등, 2007; Zheng 등, 2016b). 이스라엘에서는 B. firmus 균주를 이용한 생물학적 방제제 BipNem-WP가 시판되었으며 농장에서도 높은 살선충 효과를 나타내고 있다(Lian 등, 2007). B. firmus 배양액을 유충에 처리하면 36시간 이내에 70%의 치사율을 나타내었으며, 최근 B. firmus의 유전체 분석을 통하여 살선충 메커니즘과 병원성 인자에 대한 연구가 진행되고 있다(Zheng 등, 2016a). Bacillus속의 살선충능에는 다양한 병원성 인자들이 관련되어 있다. B. firmus의 유전체에는 B. nematocida의 단백질분해효소 Bace16, B. thuringiensis의 금속단백질분해효소 Bmp1 등 기존 보고된 Bacillus spp.와 유사한 병원성 인자를 가지고 있음이 확인되었다. B. firmus의 단백질분해효소 Sep1은 선충의 큐티클을 분해하는 병원성 인자로 밝혀졌으며, 500 μg/ml Sep1을 뿌리혹선충 유충에 처리하였을 때 58%의 치사율을 나타내는 것으로 확인되었다. 단백질분해효소 Sep1은 선충의 표피뿐만 아니라 구침과 식도 조직을 파괴하고 이후 내장 상피 세포를 분해한다. B. firmus를 생물학적 살선충제로 사용하기 위해서는 Sep1과 관련된 다양한 조절인자를 밝혀냄과 동시에 Sep1 생성량을 증가시키는 연구가 필요하다. 또한 B. firmus에 대한 생태적 안전성 및 인체 안전성에 대한 연구가 이뤄진다면 이상적인 생물학적 방제제가 될 가능성이 높다.
B. cereus: B. cereus는 1887년 처음 동정된 이후, 주로 식물병원균의 생물학적 방제균으로 많이 보고되었으며, 최근에는 살선충 활성에 대해 주목하고 있다. 명주잠자리에서 분리한 B. cereus의 sphingomyelinase C는 살충 활성이 있는 것으로 보고되었으며(Nishiwaki 등, 2004), 해성토에서 분리된 B. cereus의 경우 신규 Cry 독소인 Cry4Aa를 생성하는 것으로 보고되었다(Poopathi 등, 2014). B. cereus S2의 배양 상등액을 뿌리혹선충 유충에 처리하면 90%의 치사율을 나타내는 것으로 보고되었으며, B. cereus S2는 spinganine과 phytospingsine을 생성하여 뿌리혹선충의 활성 산소를 증가시켜 생식기관을 파괴해 번식할 수 없도록 한다(Gao 등, 2016). 뿌리혹선충 유충에 sphingosine 0.64 μg/ml를 처리하면 96시간 후 95% 치사율을 나타내었다. 또한 B. cereus를 직접 처리 시 고온과 산성조건에서 안정적이며 식물체에 뿌리혹선충에 대한 유도저항성을 일으키고, 뿌리혹선충 저항성과 관련된 유전자의 발현이 증가하는 것이 확인되었다. 그러나 B. cereus는 일반적으로는 비병원성 세균이지만 식물성 식품에 오염되었을 경우 때로는 식중독을 일으키는 경우가 있어 인체 안전성에 관한 추가적인 연구가 필요하다.
B. pumilus: B. pumilus는 1901년 처음 동정되었으며, 항진균물질, 생물계면활성제, 분해효소를 생산하여 식물병원균의 생장을 억제한다고 알려져 있다(Ahmadian 등, 2007; Akbulut 등, 2013). B. pumilus를 처리하면 고구마뿌리혹선충의 뿌리혹 형성과 알 부화가 억제되며(Akhtar와 Siddiqui, 2008; Ramezani 등, 2014), 다양한 Meloidogyne속 선충에 대해 살선충 활성을 나타내는 것으로 보고되었다(Oliveira 등, 2007). B. pumilus는 강한 키틴분해효소와 단백질분해효소 활성을 나타내며 유충의 표면과 난각을 분해하는 것으로 관찰되었다. B. pumilus의 효소 추출액 0.07 mg/ml에 79%의 치사율을 나타내며 5일 뒤에는 유충을 98%까지 치사시킨다. 또한 온실에서의 실증 실험에서도 처리 6주 후까지 뿌리혹 형성과 알 부화를 60% 이상 감소시켜 강한 살선충 활성을 확인했지만(Lee와 Kim, 2016) B. pumilus의 병원성 인자인 효소가 선충에 작용하는 메커니즘에 대해서는 아직 연구가 미비하다.
Brevibacillus laterosporus: B. laterosporus는 1916년에 Bacillus laterosporus로 동정되었다가 1996년 Brevibacillus속으로 재분류되었으며, 대두시스트선충, 소나무재선충, 동물기생선충(Trichostrongylus colubriformis), Panagrellus redivivus 등의 선충에 기생하는 기회기생균이다(de Oliveira 등, 2004; Huang 등, 2005a). 선충 표면에 정착한 뒤 선충 표피의 큐티클에 침투하여 표피 조직이 분해된다(Decraemer 등, 2003; Huang 등, 2005a). 결국 B. laterosporus가 선충 내부로 침투하면 선충 내부 조직을 분해하고 영양분으로 이용한다(Huang 등, 2005a). B. laterosporus가 분비하는 분해효소 중, 염기성 세린 단백질분해효소 BLG4가 병원성 인자로 확인되었다(Tian 등, 2006).
Fictibacillus phosphorivorans: 그람-양성 세균으로 내생포자를 생성하는 F. phosphorivorans는 2013년 생물반응기에서 분리된 이후로 갯벌과 땅콩 근권과 같은 식물 주위에서 분리되었으며, 인을 제거하여 토양 정화에 응용할 수 있다(Glaeser 등, 2013). F. phosphorivorans는 예쁜꼬마선충과 뿌리혹선충에 살선충 활성을 나타내어 유전체 분석을 통해 병원성 인자를 탐색하는 연구가 진행되고 있다. F. phosphorivorans의 유전체 내에는 amidophos-phoribosyl transferase PurL (Xia 등, 2011), 염기성 세린 단백질분해효소 BLG4 (Huang 등, 2005a), calcium-transporting ATPase Eca1 (Fan 등, 2007) 등 9개의 병원성 인자로 예상되는 유전자가 발견되었다. F. phosphorivorans는 병원성 인자만 밝혀졌을 뿐 아직 병원성 인자의 특성과 살선충 작용 메커니즘에 대해서는 연구가 미비하여, 생물학적 방제제로 이용하기 위한 실증 연구가 필요하다.
④ 방선균(Sterptomycetes)
방선균은 독소, 분해효소 등의 살선충 활성을 가진 물질을 생성한다(Rashad 등, 2015; Ruanpanun 등, 2011; Sun 등, 2006; Yang 등, 2013; Zeng 등, 2013). S. avermitilis가 만들어내는 에버멕틴은 옥수수, 목화, 콩 등 다양한 작물의 식물기생성 선충을 방제하기 위하여 AVICTA®라는 제품명으로 상품화되었다(Cabrera 등, 2013). S. avermitilis 이외의 살선충 활성을 나타내는 방선균도 생물학적 선충 방제제로서 친환경 농업에 유용하게 쓰일 수 있으리라 기대된다.
S. hydrogenase: S. hydrogenase DH16은 기존 보고되었던 방선균들과 유사하게 식물병원균이나 해충의 공격을 억제하는 것으로 이미 보고된 바가 있다(Kaur와 Manhas, 2014; Kaur 등, 2014). S. hydrogenase DH16으로부터 분리한 살선충 물질 SH2는 10-(2, 2-dimethyl-cyclohexyl)-6, 9-dihydroxy-4, 9-dimethyl-dec-2-enoic acid methyl ester로 동정되었다(Fig. 2B). S. hydrogenase가 생산하는 SH2를 200 μg/ml 처리 시 72시간 후에 유충을 100% 치사하는 것으로 확인되었다(Kaur 등, 2016). S. hydrogenase DH16은 살선충 활성에 대한 실증실험 결과가 미비하고, 산업적 적용을 위해서는 대량 배양과 제제화 연구가 필요하다.
Streptomyces spp.: Streptomyces sp. CMU-M021이 생산하는 fervenuline과 isocoumarin은 진균에 길항능을 나타내며 선충을 치사시킨다(Fig. 2B). Fervenuline은 배양 후 최대 160시간 동안에 유충은 100% 치사하고, 알 부화율은 5%로 감소했다. Isocoumarin의 경우 250 μg/ml 농도로 처리 시 유충을 50% 치사하였다(Ruanpanun 등, 2011). 이와 유사하게 S. albogriseolus가 생산하는 항진균 물질인 fungichromin B도 고구마뿌리혹선충에 대하여 15 μg/ml를 처리했을 때 100% 치사하는 것으로 보고되었다(Fig. 2B; Zeng 등, 2013).
⑤ 기타 살선충 활성을 나타내는 세균
Serratia mercescens: 프로디지오신(prodigiosin)은 적색 색소 물질로 S. marcescens가 만드는 이차 대사 산물이며, Streptomyces coelicolor도 생성하는 것으로 보고되었다(Williamson 등, 2006). S. marcescens는 항생제, 면역억제제, 항암제를 포함한 생리활성능을 가지고 있는 것으로 알려졌다. 프로디지오신을 160 μg/ml 처리 시 뿌리혹선충, 감귤선충의 유충에 대하여 100%의 치사율을 나타내었고, 알 부화도 억제하는 것으로 보고되었다(Suryawanshi 등, 2014). 그러나 프로디지오신의 살선충 메커니즘은 아직 밝혀지지 않았다.
Alcaligenes faecalis: 1919년, 배설물에서 처음 발견되었으며 토양, 지하수, 식물, 선충 등 다양한 환경에 서식하고 있다(Phung 등, 2012; Rehfuss와 Urban, 2005). A. faecalis는 다양한 분해효소를 생성하여 유기오염물질을 분해하기 때문에 폐수정화 등에 이용되고 있으며(Kong 등, 2014; Rehfuss와 Urban, 2005) 만델산, 다양한 의약품의 전구체를 생성하는 것으로 알려져 의료산업에도 적용되고 있다(Kong 등, 2014; Xia, 2013; Yamamoto 등, 1991). A. faecalis 균주는 56시간 이내에 뿌리혹선충 유충을 65% 이상 치사하는 것으로 보고되었다. 그러나 80°C에서 열처리를 하였을 때는 치사율이 50%로 감소하였다. A. faecalis의 유전체 분석 결과, 세린 단백질분해효소인 Esp를 확인하였으며, 400 μg/ml Esp 처리 시 뿌리혹선충 유충을 100% 치사하여 병원성 인자임이 밝혀졌으며, 선충 내부로 침투하고 24시간 이내에 장 조직을 파괴하여 선충을 치사시키는 것으로 알려졌다(Ju 등, 2016). 아직 분자적 메커니즘과 온실 효과 실증 실험에 대한 연구가 미비하지만, 토양에서 정착이 가능하여 근권토양에 우점세균으로 존재할 수 있다는 점 때문에 생물학적 방제제로 개발하였을 때 높은 지속력이 기대된다.
진균에 의한 선충의 생물학적 방제 방법
선충포식성 진균은 토양 내에서 선충과 천적관계를 이루고 있는 진균으로 기후를 가리지 않고 지구상의 다양한 환경에 존재하고 있다(Nordbring-Hertz 등, 2011). 이들은 특수한 구조를 이용하여 살아있는 선충을 공격하는 선충포획 진균, 균사의 끝으로 선충 내부와 알에 침투하는 내부기생 진균, 알기생 진균, 선충을 공격하기 전 독소를 분비하여 움직이지 못하게 하는 독소 생성 진균로 분류할 수 있다(Fig. 4; Liu 등, 2009; Moosavi와 Zare, 2012; Yang과 Zhang, 2014). 선충포식성 진균은 자낭균, 담자균, 접합균, 병꼴균 등에 속해 있으며 700종 이상 보고되었다(Yu 등, 2014). 실제로 몇몇 선충포식성 진균의 경우 스위스, 독일, 오스트리아 등지에서 생물학적 방제제로 사용되고 있다(BLW, 2015; BVL, 2015; Degenkolb and Vilcinskas, 2016).
Fig. 4
Pathogenesis of nematophagous fungi. On the basis of the mechanisms that fungi use to infect nematodes, nematophagous fungi can be categorized into three major groups: nematode-trapping fungi (A), endo-parasitic fungi (B), and egg-parasitic fungi (C). Nematode-trapping fungi share a unique ability to form specialized trapping structures (traps) to capture nematodes. Endo-parasitic fungi are a group of fungi that use their conidia or zoospores to invade inside nematodes. Egg-parasitic fungi use appressoria, a specialized penetration peg, or lateral mycelial branches to infect nematode eggshells.
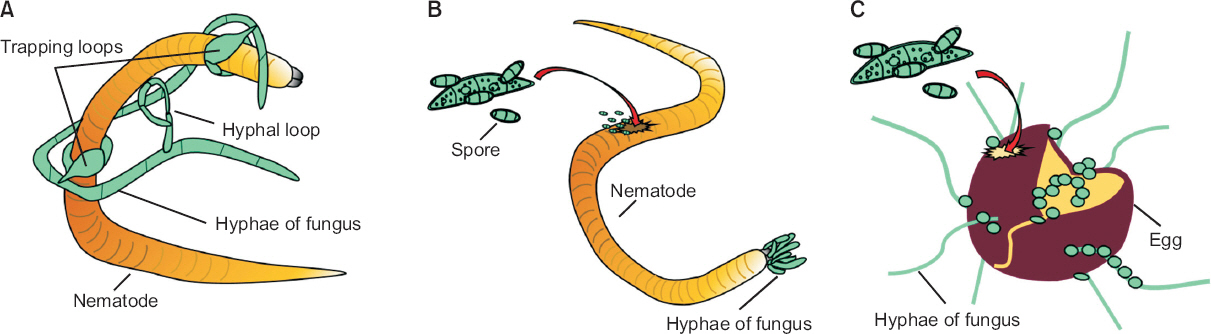
① 선충 포획 진균
선충포획 진균은 특수한 형태의 포획구조를 생성하여 살아있는 선충의 유충, 성충, 알을 공격해 영양원으로 사용한다. 지금까지 알려져 있는 선충포획 진균은 대부분 자낭균문의 바퀴버섯목에 속해 있다(Yang 등, 2007b, 2012). 일반적으로 널리 알려진 진균은 Arthrobotrys (54종), Drechslerella (14종), Dactylellina (28종)속이다(Yu 등, 2014). 이들은 절대적 포식균이 아니기 때문에 선충이 존재하지 않을 때에는 토양 내에서 부생균으로 남아 있다.
선충포획 진균은 포획구조의 형태에 따라 점착성 그물구조를 형성하는 Arthrobotrys oligospora, 점착성 균사를 만드는 Dactylellina haptotyla, 선충을 가두는 링 구조를 만들어내는 Drechslerella stenobrocha의 3가지 형태로 분류되었다(Liu 등, 2014; Meerupati 등, 2013; Yang 등, 2011). 선충포획 진균 중에서 A. oligospora, D. haptotyla, D. stenobrocha의 유전체 정보가 밝혀져 포획구조 형성 메커니즘에 대한 연구가 활발하게 이루어지고 있다(Liu 등, 2014; Meerupati 등, 2013; Yang 등, 2011). 선충포획 진균의 포획구조 형성을 조절하는 신호경로와 핵심 유전자에 대한 연구는 현재 매우 흥미로운 주제이다.
Arthrobotrys spp.: 대표적인 선충 포획 진균으로 Arthrobotrys속의 A. oligospora, A. dactyloides, A. conoides가 보고되고 있다. A. oligospora는 다른 선충포획 진균에 비해 증식속도가 빨라 균사 성장과 포자의 발아가 쉽고, 국내뿐만 아니라 전 세계에서 삼림, 농경지, 퇴비더미 등 거의 모든 지역에 분포한다(Yang 등, 2011). 이들은 A. oligospora, A. conoides의 포획기관은 모두 3차원적 점착성 입체적 그물구조의 형태로 일반 균사가 곁가지를 내고 그것이 구부려져 고리모양을 이룬다. 이 고리모양이 이웃하는 다른 고리와 합쳐져 더 큰 고리를 만들고 수많은 고리들이 전체적으로 서로 얽혀 입체적인 망 구조를 형성하기 때문에 선충이 잡히면 빠져 나오지 못한다(Meerupati 등, 2013). A. dactyloides는 압축성 고리 형태로 완전한 원의 형태를 띠며 고리끼리 합쳐지지 않는다. 이 압축고리는 3개의 세포로 이루어져 있으며 이 세포가 팽창하여 고리 안에 들어온 선충을 조인다. 이 진균들은 선충의 움직임이나 분비물 등을 인식하기 시작하면 수분 내에 포획기관을 형성한다. 그리고 이 포획기관에 선충이 잡히게 되면 더 많은 접착 물질을 분비하여 선충 표면의 분비물과 화학적 결합을 일으킨다(Meerupati 등, 2013).
포획 진균이 선충을 인지하면 포획기관은 빠르게 부착단백질을 만들어 숙주인 선충에 점착한다(Tucker와 Talbot, 2001). 포획 진균이 선충과 접촉하면 선충 표면의 렉틴을 인지하고 점착물질을 분비한다. 렉틴은 탄수화물과 결합능력이 있는 당단백질로 포획기관의 다당류와 공유 결합하여 점착성이 더 강해진다. 점착물질이 만들어지면 포획기관의 균사 표면을 구성하는 미소섬유 중합체(extracellular fibrillar polymers)의 밀도가 높아져 선충과 부착하게 되고 이후 여러 가지 단백질 분해효소가 선충을 분해한다(Li 등, 2015).
일반적으로 선충포획 진균은 침입도구를 형성하기 위해 선충이 만들어내는 물질을 인식한다고 추정하고 있는데 A. conoides는 선충이 생성하는 네민이라는 물질을 인지하여 포획구조를 형성한다는 연구결과가 발표되었고(Pramer와 Stoll, 1959), A. oligospora와 근연종들 또한 선충이 분비하는 저분자 물질을 인식하여 포획구조를 형성함이 보고되었다(Hsueh 등, 2013). 이러한 연구들을 통하여 선충 포획 진균은 선충과 화학적 상호작용을 통하여 인식하고 포획구조를 형성함이 알려졌다.
A. dactyloides가 포획구조를 형성하기 위해서는 G-단백질 신호전달체계가 있어야 한다(Chen 등, 2001). G-protein은 세포 내의 칼슘 농도를 증가시켜 고리구조가 부풀도록 활성화시키거나, 반대로 부풀지 않도록 비활성화시키기도 한다고 알려졌다. A. oligospora에서도 렉틴과 결합하는 GLEYA (a conserved motif G (M/L) (E/A/N/Q)YA) 도메인과 CFEM 도메인(common in several fungal extracellular membrane proteins)을 포함한 단백질이 분리되었는데(Kulkarni 등, 2005; Yang 등, 2011), 이 단백질들은 D. stenobrocha, D. haptotyla에서도 분리되었으며 선충포획 진균의 병원성에 중요한 인자일 것으로 추측되었다(Liu 등, 2014; Meerupati 등, 2013). 이 외에도 유전자 정량 분석을 통해 A. oligospora의 17개의 부착단백질과 관련된 유전자를 발견하였으며 이 중 5개 유전자는 포획구조를 형성하는 동안 발현이 증가하는 것으로 확인되었다(Yang 등, 2011). 또한 이 5개 유전자는 곤충병원성 진균인 M. anisopliae가 곤충 표면에 부착하는 것을 조절하는 것으로 알려진 MAD1과 상동유전자인 것으로 확인되었다(Wang과 St. Leger, 2007). 선행연구들을 통하여, 선충포획 진균의 포획구조 형성을 조절하는 유전자와 단백질이 밝혀졌지만, 자세한 조절 메커니즘과 이후 과정들에 대한 정보는 알려지지 않았다.
② 선충 내부 기생 진균
선충 내부 기생 진균은 분생자 또는 포자를 이용하여 선충에 감염한다. 내부 기생 진균은 선충을 인지하면 빠르게 분생자를 형성하고 균사를 이용하여 선충 내부에 침투한다. 기생성 진균은 포획성 진균과는 달리 대부분 균사체를 형성하지 않아 선충이 없으면 토양에서 증식하기 어렵다고 알려져 있었지만 선충이 없을 때에는 토양 내에서 식물의 뿌리 분비물이나 죽은 뿌리 부분을 양분으로 하여 생존하기도 하기 때문에 토양에 정착이 가능한 생물학적 방제제로 이용이 가능하다(Moosavi와 Zare, 2012).
Drechmeria coniospora: D. coniospora의 경우 선충 1마리에 10,000개의 포자를 형성하여 감염하는 것으로 알려져 있다. 선충 내부에 침투한 후, 큐티클 층에서 소포가 감염되면 내부에서 균사체가 형성되어 3일 내에 선충을 분해한다(Moosavi와 Zare, 2012). D. coniospora의 포자가 성장을 마치면 한 쪽 끝에 점착성의 혹을 만들어 선충의 큐티클이나 구강, 생식기에 부착한다. 부착한 후, 선충 내부에서 감염성의 소포를 만들어 영양균사를 만들기 시작한다. D. coniospora의 균사가 자라면서 3일 내에 선충을 분해하고 죽은 선충 안에서 분생자병이 만든 포자들이 방출된다(Moosavi와 Zare, 2012). 최근 D. coniospora의 유전체가 분석되어(Lebrigand 등, 2016), 2차대사 조절인자 LaeA, 다양한 단백질분해효소, 이열성 장내독소(heat-labile enterotoxin) 생산 유전자 등 병원성 인자로 예상되는 11개의 유전자가 발견되었다. 그러나 D. coniospora의 진균-숙주 간 상호작용에 대한 연구는 아직 미비하다.
Esteya vermicola: E. vermicola는 타이완에서 최초로 발견된 선충 내부 기생 진균이다(Liou 등, 1999). 아시아, 유럽, 남아메리카 등 전 세계적으로 분포하며 나무좀(Scolytus intricatus)에 의해 옮겨지고 나무 안에서 부생균으로 존재하면서 소나무재선충에 감염한다(Kubátová 등, 2000; Liou 등, 1999; Wang 등, 2008, 2014). E. vermicola의 기주 범위는 소나무재선충뿐만 아니라 벼이삭선충(Aphelenchoides besseyi), 딸기잎선충(A. fragariae), 국화잎선충(A. ritzemabosi), 벼줄기선충(Ditylenchus angustus), 마늘구근선충, 감자썩이선충(D. destructor), red ring nematode (Rhadinaphelenchus cocophilus), 고구마뿌리혹선충, 귀리시스트선충(Heterodera avenae), 뿌리썩이선충(Pratylenchus penetrans) 등이 포함된다(Kubátová 등, 2000; Wang 등, 2008, 2014).
E. vermicola는 두 가지 종류의 분생자의 생성하는 것으로 알려져 있는데 이 중 루네이트(lunate)만이 점착성을 나타내어 선충 큐티클층에 부착하고 선충에 감염한다(Liou 등, 1999). 루네이트가 선충 표면에 부착하면 18-24시간 안에 분생자가 발아하고 선충 표면의 큐티클에 부착기를 형성하여 내부로 균사를 만들기 시작한다. 부착된 분생자의 수와 발아 속도에 따라 2-3일 내에 균사가 자라나 선충의 기관과 조직을 파괴하여 치사한다. 내부 기생 진균은 포자에서 비휘발성 삼출물과 휘발성 물질로 된 화학물질을 내뿜어 멀리서부터 선충을 유인하는 것으로 알려져 있다(Chu 등, 2015). E. vermicola가 생산하는 휘발성 화합물의 조성을 분석한 결과, α-pinene, β-pinene, camphor (28:12:1)로 구성되어 있음이 보고되었다(Fig. 2B; Lin 등, 2013). 이 휘발성 화합물들은 소나무와 나무좀에서도 만들어지며 소나무재선충을 유인하는 것으로 보고되었다(Lin 등, 2013). E. vermicola가 이 화합물을 이용하여 어떻게 선충을 유인하는지에 대한 연구가 필요하다. 또한 E. vermicola의 병원성 인자로써 키틴분해효소 유전자가 보고되기는 했지만 아직 키틴분해효소의 특성과 메커니즘에 관해서는 알려지지 않았다(Wei 등, 2014). E. vermicola는 저온에서 배양했을 때 태양열이나 건조 등 다양한 환경스트레스에 내성을 얻고 소나무재선충에 대한 감염성이 늘어나 실제 재배지 적용 후에 지속성이 향상되는 것으로 보고되었다(Wang 등, 2012). 아직 E. vermicola가 선충을 감염하는 메커니즘, 진균-숙주 간 상호작용에 대한 정보는 미비하고 실증 실험 모니터링과 제품화에 대한 연구가 필요하다.
Myrothecium verrucaria: M. verrucaria는 Tuberculariaceae과에 속하는 불완전균류로 주로 식물과 토양에서 분리되었고(Tulloch, 1972), 최근까지 생물제초제로서 주목 받아왔다(Abbas 등, 2001; Walker와 Tilley, 1997; Yang 등, 1991). M. verrucaria의 배양액을 고구마뿌리혹선충에 사용했을 때 선충을 치사시키고 알 부화율이 감소하는 것으로 나타났으며, 감자몬스트포낭선충과 마늘구근선충의 기주식물 인식과 이동을 억제하는 것으로 보고되었다(Fernández 등, 2001; Twomey 등, 2000). 또한 M. verrucaria는 당근뿌리혹선충의 알과 유충, 성충에 기생하는 것으로 보고되었다(Dong 등, 2015). M. verrucaria는 난각 주변을 균사로 둘러싸고 키틴분해효소를 생성하여 파괴하며, 유충을 마비시키고 분생자가 내부로 침투하여 균사를 성장시키고 단백질 분해효소를 생성하여 유충을 분해한다. 성충의 경우 표면을 균사가 그물처럼 둘러싼 후 단백질 분해효소를 생성하여 분해한다. M. verrucaria는 당근뿌리혹선충의 알과 유충, 성충에 감염하여 107 conidia/ml로 처리 시 3일 이내에 70% 이상의 살선충 활성을 나타내었다. 또한 뿌리혹선충에 감염된 오이 재배지에서도 M. verrucaria 처리 시 유충의 수가 감소하는 것으로 보고되었다(Dong 등, 2015). 그러나 M. verrucaria의 살선충 활성을 나타내는 병원성 인자와 작용 메커니즘에 대한 연구는 아직 미비하다. M. verrucaria는 DiTera®라는 이름으로 제품화되어 선충 방제를 위한 대표적인 생물학적 방제제 중 하나로 알려져 있으며 채소 및 과일 재배지의 뿌리혹선충, 시스트선충, 뿌리썩이선충에 방제에 사용할 수 있다(Wilson과 Jackson, 2013).
③ 선충 알 기생 진균
알 기생 진균은 특수한 구조의 부착기를 사용하거나 균사가 측면으로 분기하여 선충의 난각(eggshell)에 감염한다(Lopez-Llorca 등, 2008). 알 기생 진균이 생성하는 키틴분해효소, 젤라틴분해효소는 키틴과 단백질로 구성된 선충의 알 표면을 분해한다(Yang 등, 2007a). 알 기생 진균은 맥각균과에 속해 있는 Pochonia chlamydosporia, Purpureocillium lilacinus, Clonostachys rosea, Lecanicillium psalliotae로 곤충병원성 진균과 분류학적으로 매우 가깝다. 최근 P. chlamydosporia의 염기서열이 분석되어(Larriba 등, 2014) 영양조건에 따른 진균의 발달 연구에 많은 도움이 될 것으로 생각된다.
Pochonia spp.: P. rubescens와 P. chlamydosporia는 식물 뿌리 내에서 존재하다가 식물기생성 선충의 알에 감염한다. 1990년 최초로 P. rubescens에서 선충 표면의 큐티클을 분해하는 세린 단백질분해효소 P32가 처음으로 동정되었고(Lopez-Llorca, 1990), 근연종인 P. chlamydosporia의 단백질분해효소 VCP1도 보고되었다(Segers 등, 1994). 또한 P. rubescens는 선충의 알 표면에 침투하기 위하여 키틴분해효소 Chi43을 이용하는 것으로 보고되었다(Tikhonov 등, 2002). 키틴분해효소 Chi43은 P. rubescens가 선충 알에 기생하기 위한 병원성 인자로 밝혀졌고, 감자시스트선충(Globodera pallida)의 알 표면에 키틴분해효소 Chi43과 단백질분해효소 P32를 처리 시 알 표면에 상처를 내어 껍질이 벗겨지는 것을 전자현미경(scanning electron microscope)을 통해 관찰하였다(Tikhonov 등, 2002). 앞으로 P. rubescens와 P. chlamydosporia의 세포 외 분해효소의 특성 및 메커니즘을 규명하는 연구가 필요할 것으로 보인다.
Purpureocillium lilacinus: P. lilacinus는 다양한 식물기생성 선충 알에 기생하는 것으로 알려져 P. lilacinus 251의 경우 이미 생물학적 방제제로 제품화되어 있다(Brand 등, 2004). P. lilacinus는 선충의 알에 특이적으로 기생하며, 다양한 작물에 감염된 뿌리혹선충과 감자몬스트포낭선충을 효과적으로 방제한다고 보고되었다(Khan 등, 2006). P. lilacinus는 선충을 인식한 후 12시간 안에 포자가 발아하고 부착기를 만들어 선충 내부로 침투한 뒤 24시간 내에 선충 알을 파괴한다(Kiewnick와 Sikora, 2006). P. lilacinus는 감염 과정 동안에 류시노톡신(leucinotoxin), 키틴분해효소, 큐티클분해효소, 단백질분해효소, 아세트산을 생성하는 것으로 보고되었다(Park 등, 2004). 이 중 키틴분해효소와 단백질분해효소가 뿌리혹선충에 대해 중요한 병원성 인자로 작용하며, 키틴분해효소가 난각 구조를 변형시켜 단백질분해효소가 작용할 수 있도록 보조한다는 연구결과가 보고되었다(Khan 등, 2004). 현재 P. lilacinus의 분해효소의 특성 및 메커니즘에 대한 연구가 진행 중이다(Khan 등, 2003). P. lilacinus를 생물학적 제제로 이용하기 위해서는 제형화 및 배양 조건 확립 등의 연구가 필요하다.
④ 독소 생성 진균
독소 생성 진균은 선충의 큐티클에 침투하기 전에 독소를 분비하여 선충을 마비시켜 움직이지 못하게 한다(Lopez-Llorca 등, 2008). 자낭균류와 담자균류에 속한 280여 종의 진균에서 살선충 활성을 가진 독소 200여 개가 분류되었으며, 독소 물질들은 알칼로이드, 펩타이드, 테르페노이드, 마크로라이드, 벤젠 화합물, 퀴논, 지방족화합물, 단순방향족화합물로 이루어져 있다(Li와 Zhang, 2014; Li 등, 2007a).
Talaromyces thermophilus: 최근 T. thermophilus가 생성하는 써모라이드(thermolides)는 상업적으로 판매되고 있는 에버멕틴과 유사할 정도로 강한 살선충능을 나타내는 것으로 발표되었다(Guo 등, 2012). 써모라이드를 이용하여 선충을 방제하기 위해 실증실험과 안전성 평가 등 생물학적 방제제 개발을 위한 연구가 활발히 이루어지고 있다.
Coprinus comatus: C. comatus는 독소를 형성하면서 동시에 spiny ball, acanthocyte라는 유인구조를 생성한다(Luo 등, 2007). 이 유인구조는 선충의 큐티클에 상처를 내어 치사하게 만드는 병원성 인자로 보고되었다. C. comatus는 7가지의 독소를 생성하는 것으로 알려져 있는데, 이들은 모두 퓨라논(furanone)과 유사한 구조를 가지는 것으로 보고되었다. 이 중 5-ethylfuran-3-carboxylic acid, 5-Hydroxy-3,5-dimethylfuran-2(5H)-one은 고구마뿌리혹선충 유충에 200 μg/ml 농도로 처리 시 90% 이상의 높은 살선충 활성을 나타내었다(Fig. 2B; Luo 등, 2007). C. comatus가 생성하는 독소를 생물학적 제제로 이용하기 위해서는 작물 및 동물에 대한 독성평가가 필요하다.
⑤ 기타 살선충 활성을 나타내는 진균
4가지의 포식성 진균들 외에도 다양한 진균들이 살선충 활성을 가지고 있다(Fig. 2B, Fig. 2 Table 2). Tricoderma spp.는 선충의 알과 유충에 기생하면서 단백질분해효소 PRA1, SprT, 키틴분해효소 chi18-5, chi18-12와 같은 다양한 분해효소들을 분비하는 것으로 알려졌다(Suarez 등, 2004; Szabó 등, 2012; Vachon 등, 2012). 또한 trichodermin, β-vinylcyclopentane-1α, 3α-diol, 6-pentyl-2H-pyran-2-one, 4-(2-hydroxyethyl) phenol 등의 살선충 활성을 나타내는 독소도 생성하는 것이 확인되었다(Fig. 2B; Yang 등, 2010). 균근은 선충에 감염된 식물의 생장을 향상시키고 저항성을 유도하여 간접적으로 선충 피해를 감소시키는 것으로 보고되었다(Schouteden 등, 2015; Veresoglou와 Rillig, 2012; Vos 등, 2012). 누룩곰팡이로 알려져 있는 Aspergillus속 진균도 살선충 활성을 나타내는 것으로 보고되었다. A. oryzae가 생산하는 누룩산(kojic acid)과 A. niger가 생산하는 옥살산이 살선충활성을 나타내는 것으로 보고되었다(Jang 등, 2016; Kim 등, 2016). A. niger가 생산하는 옥살산의 경우 2 μM/ml 처리 시 고구마뿌리혹선충 유충을 24시간 이내에 100% 치사시키고 알 부화를 7일 동안 95% 이상 억제하는 것으로 보고되었다. 또한 온실실험에서 뿌리혹형성이 대조구에 비하여 60% 이상 감소하는 것으로 나타났다. A. niger의 경우, 옥살산과 A. niger를 혼합한 제품이 뿌리혹선충을 방제하기 위한 생물학적 방제제로 국내에서 제품화되었다.
이처럼 다양한 진균들이 살선충 활성을 나타내는 것으로 확인되었으며, 이를 실제 시설 재배지에서 선충에 대한 생물학적 방제 효과를 증명하고 농가에 보급하고자 국내외에서 많은 연구들이 이루어지고 있다.
한계점: 최근까지 개발된 미생물 제제들의 경우 실내 검정에서 우수한 효과를 나타내었으나 실재 재배지에 처리 시 효과와 안정성이 화학적 방제제에 미치지 못하는 것으로 알려졌다. 이에 따라 안정적인 효과를 나타내는 생물학적 선충 방제제의 개발이 시급하다. 살선충 활성을 나타내는 미생물과 토양 개량제를 혼합하여 처리 시 미생물 단일 처리보다 살선충 활성이 증가하였다는 보고가 이어져, 선충 방제 미생물과 혼합하여 사용한다면 식물기생성 선충을 방제효과와 지속성을 향상시키는 방법이라고 생각된다. 또한 식물기생성 선충의 방제 시점과 처리 방법을 최적화하고, 효과 증진을 위해 대량배양 생산과 토양 정착을 위한 제형화 기술 개발이 필요할 것이다. 지금까지 살선충 활성을 나타내는 미생물들이 보고되었고, 식물기생성 선충을 방제하기 위한 생물학적 방제제로서 개발되었지만 살선충 활성과 메커니즘, 살선충 물질의 합성 경로에 대한 정보는 부족하다. 새로운 살선충 물질에 대한 합성 경로, 작용 메커니즘, 실증효과, 선충-미생물 간의 상호작용에 대한 연구가 수행된다면 안정적이고 높은 효과를 나타내는 생물학적 선충 방제제를 개발할 수 있을 것이다.
결언
지금까지 식물병원성 해충과 선충에 대해 살충/살선충 활성을 나타내는 세균, 진균, 바이러스를 소개하고 대표적인 살충/살선충 활성 물질을 살펴보았다(Table 1, 2, Fig. 1⁻4). 방제제 개발 가능성이 있는 것으로 알려진 다양한 미생물들이 최근 연구를 통하여 병원성 인자, 작용 메커니즘이 밝혀지고 있어 앞으로 개발해서 적용할 가능성이 높다. 실제 재배지 적용을 위해서는 미생물 정착능을 높이고 배양조건을 최적화하여 제형화 기술이 개발되어야 할 것이다. 또한 방제능을 유지할 수 있는 안정화 기술을 확보하고 인체 안전성을 확보한다면 화학적 방제제를 대체하여 사용률이 높아질 것이라고 생각된다. 방제능을 가진 다양한 미생물의 제품 개발 기술을 발전시킨다면 차기 친환경 농업의 유망 기술로 주목 받을 수 있을 것이다.
요약
전 세계적으로 주요 작물에서 식물 병원성 해충 및 선충으로 인한 작물 생산량 감소 등의 경제적 손실이 심각하다. 우리나라의 경우 시설 재배지의 연작으로 인해 해충과 선충의 발생이 증가하여 작물의 피해가 점차 증가하고 있는 추세다. 현재 주로 사용되고 있는 식물병원성 해충 및 선충의 방제법은 사용이 편리하고 비용이 저렴한 화학적 방제법을 이용하고 있지만 화학 농약의 남용으로 인한 약제 저항성이 발생하고, 화학 약품으로 인한 인체 유해성 및 환경오염 문제가 대두되고 있다. 이로 인해 최근 친환경적인 방제에 관한 지속적인 연구와 관심이 높아지면서 식물병원성 해충과 선충에 대하여 살충/살선충 활성을 나타내는 미생물을 생물학적 방제법으로 이용하기 위한 시도가 증가하고 있다. 본 리뷰에서는 식물병원성 해충과 선충에 대한 살충/살선충 활성을 나타내는 세균, 진균, 바이러스에 의한 살충 및 살선충 사례와 기전들을 소개하였다. 이러한 미생물의 활성은 외피 분해효소와 독소물질생산을 통한 직접적인 작용기전과 기주 식물의 저항성 유도를 통한 간접적인 작용기전에 의해서 일어난다. 본 리뷰를 통하여 선충과 해충에 대한 생물학적 방제법과 그 분자 작용기전에 대한 이해를 돕고, 최신 살충 및 살선충 연구들을 소개함으로써 국내의 농민들과 연구자들의 미생물을 기반으로 한 친환경적 해충 및 선충의 방제법에 대한 저변확대를 기대한다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






