

ņä£ļĪĀ
Ļ│╝ņłś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ(black shoot blight)ņØĆ 1995ļģä Ļ░ĢņøÉļÅä ņČśņ▓£ĻĘ╝ĻĄÉņØś ļ░░(Pyrus pyrifolia cv. ŌĆśShingoŌĆÖ) Ļ│╝ņøÉņŚÉņä£ ņĄ£ņ┤łļĪ£ Ļ┤Ćņ░░ļÉśņŚłļŗż. ļ│æņ¦ĢņØĆ ņŚĮļ®┤ņØś ĒØæĻ░łņāē ļ░śņĀÉĒśĢņä▒Ļ│╝ Ļ┤┤ņé¼ņ”Øņāü, ņ×ÄņØś ņŻ╝ļ¦źņØä ļö░ļØ╝ ĒØæĻ░łņāē ņżäļ¼┤ļŖ¼Ļ░Ć ĒśĢņä▒ļÉśņŚłņ£╝ļ®░, ņØ┤ļ¤¼ĒĢ£ ņ”ØņāüņØĆ ņżäĻĖ░ņĀäņ▓┤ļĪ£ ĒÖĢņé░ļÉśņ¢┤ Ļ░£ĒÖöĻĖ░ņÖĆ ņ░®Ļ│╝ĻĖ░ņŚÉļŖö Ļ░üĻ░ü ĻĮāĻ│╝ ļ»Ėņä▒ņłÖĻ│╝ņŚÉ ļ¦łļ”ä ļ░Å Ļ┤┤ņé¼ņ”ØņāüņØ┤ ļéśĒāĆļé¼ļŗż. ņØ┤ļŖö ņé¼Ļ│╝ ļ░Å ļ░░ ĒÖöņāüļ│æ(fire blight)ņØś ļ│æņ¦ĢĻ│╝ ļ¦żņÜ░ ņ£Āņé¼ĒĢśņŚ¼, ļŗ╣ņŗ£ņŚÉļŖö ĻĄŁļé┤ņŚÉņä£ ĒÖöņāüļ│æņØ┤ ļ░£ņāØĒĢ£ Ļ▓āņ£╝ļĪ£ ņśżņØĖļÉśņ¢┤ ļ¦ÄņØĆ ļģ╝ļ×ĆņØä ņĢ╝ĻĖ░ņŗ£ņ╝░ļŹś ļīĆĒæ£ņĀü ņäĖĻĘĀņä▒ Ļ│╝ņłśļ│æĒĢ┤ņØ┤ļŗż. ĻĘĖļ¤¼ļéś ļ│æņøÉņäĖĻĘĀņØś ļ░░ņ¢æņĀü, Ēśłņ▓ŁĒĢÖņĀü ļ░Å ņ£ĀņĀäĒĢÖņĀü ĒŖ╣ņä▒ņØä ļČäņäØĒĢ£ Ļ▓░Ļ│╝, ĒÖöņāüļ│æņØś ļ│æņøÉņäĖĻĘĀņØĖ Erwinia amylovoraņÖĆļŖö ĒŖ╣ņä▒ņØ┤ ņÖäņŚ░Ē׳ ļŗżļźĖ ļ│æņøÉņäĖĻĘĀņ£╝ļĪ£ ĒÖĢņØĖļÉśņŚłņ£╝ļ®░(Rhim ļō▒, 1999), 1999ļģä Kim ļō▒(1999)ņŚÉ ņØśĒĢ┤ ņāłļĪ£ņÜ┤ ņŗĀņóģņØĖ Erwinia pyrifoliaeļĪ£ ļ¬ģļ¬ģļÉśņ¢┤ ĻĄŁļé┤ņŚÉņä£ ļ░£ņāØĒĢśļŖö ņé¼Ļ│╝ ļ░Å ļ░░ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ņøÉņØĖņäĖĻĘĀņ£╝ļĪ£ Ļ│Ąņŗ£ļÉśņŚłļŗż.
ņØ┤Ēøä ļåŹļ”╝ņČĢņé░ņŗØĒÆłļČĆ ļ░Å ļåŹņ┤īņ¦äĒØźņ▓ŁņŚÉņä£ 1995ļģäļČĆĒä░ 2016ļģäĻ╣īņ¦Ć ņ¦ĆņåŹņĀüņØĖ ļ¬©ļŗłĒä░ļ¦üņØä ņŗżņŗ£ĒĢ£ Ļ▓░Ļ│╝, 2007ļģäĻ│╝ 2010ļģäņØä ņĀ£ņÖĖĒĢ£ ļ¦żļģä Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØņØ┤ ņĪ░ņé¼ļÉśņŚłļŗż(ļåŹļ”╝ņČĢņé░ņŗØĒÆłļČĆ, Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØĻ▓ĮĻ│╝ ļ░Å ņĪ░ņ╣śņé¼ĒĢŁ ņ░ĖņĪ░). ĒŖ╣Ē׳ 2016ļģäņŚÉļÅä Ļ░ĢņøÉļÅä ņČśņ▓£, ĒÖŹņ▓£ ļ░Å ĒÜĪņä▒ ĻĘĖļ”¼Ļ│Ā Ļ▓ĮĻĖ░ļÅä Ēżņ▓£ ņé¼Ļ│╝Ļ│╝ņøÉņŚÉņä£ ļ░£ņāØņØ┤ ĒÖĢņ¦äļÉśņŚłļŗż. ņØ┤ļ¤¼ĒĢ£ ņŻ╝ĻĖ░ņĀü ļ¬©ļŗłĒä░ļ¦ü ņØ┤ņÖĖ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņŚÉ ļīĆĒĢ£ ļ░£ļ│æņøÉņØĖ, ņĀäņŚ╝Ļ▓ĮļĪ£ ļ░Å ņ¢ĄņĀ£ĻĖ░ņłĀ ļō▒ņŚÉ Ļ┤ĆĒĢ£ ņŚ░ĻĄ¼ļŖö ņé¼ņŗż ņĀäļ¼┤ĒĢ£ ņŗżņĀĢņØ┤ļŗż. ļ░śļ®┤ ĒÖöņāüļ│æņØś Ļ▓ĮņÜ░ Ļ░£ĒÖöņŗ£ĻĖ░ ĒÖöļČäļ¦żĻ░£ņČ®ņŚÉ ņØśĒĢ┤ ĻĮāņØä ĒåĄĒĢ┤ ļ░£ļ│æĒĢśĻ│Ā(BayotņÖĆ Ries, 1986; Hildebrand ļō▒, 2000), ņØ┤ļ│æĻ░Ćņ¦Ć ņĀ£Ļ▒░ņŚÉ ņØśĒĢ£ ĻĄ¼ņ▓┤ņĀü Ļ┤Ćļ”¼ļ░®ņĢł ļō▒ņŚÉ Ļ┤ĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ļŗżņłś ņłśĒ¢ēļÉśņŚłļŗż(Johnson, 2000; Sundin, 2014; Wilcox, 1994).
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļŖö 2ļģäĻ░äņØś ņĀĢĻĖ░ņĀü ļ¬©ļŗłĒä░ļ¦üņØä ĒåĄĒĢ┤, Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ļ░£ņāØņś©ļÅä ļ░Å ņŗ£ĻĖ░ ĻĘ£ļ¬ģĻ│╝ ļ│æņ¦ĢņØ┤ ļéśĒāĆļéśļŖö ņØ┤ļ│æņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ļź╝ ĒÖĢņØĖĒĢśņŚ¼ ņØ┤ļ│æņŻ╝ņØś ļ¼┤ņĪ░Ļ▒┤ņĀüņØĖ ņĀ£Ļ▒░Ļ░Ć ņĢäļŗī ļ│æņøÉņäĖĻĘĀ Ļ▓ĆņČ£ ļČĆņ£äļ¦īņØä ņĀ£Ļ▒░ĒĢśļŖö ĒÜ©ņ£©ņĀü Ļ░Ćņ¦Ćņ╣śĻĖ░ ļ░®ļ▓ĢņØä ĒåĄĒĢ£ Ļ┤Ćļ”¼ļ░®ņĢłņØä ļ¦łļĀ©ĒĢśĻĖ░ ņ£äĒĢśņŚ¼ ņłśĒ¢ēļÉśņŚłļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ņØ┤ļ│æņ▓┤ ņłśņ¦æ ļ░Å ļ│æņøÉņäĖĻĘĀ ļČäļ”¼
ļåŹļ”╝ņČĢņé░ņŗØĒÆłļČĆ ŌĆ£Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØĻ▓ĮĻ│╝ ļ░Å ņĪ░ņ╣śņé¼ĒĢŁŌĆØņŚÉ ļö░ļØ╝ 2013ļģä Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØņØ┤ ņĄ£ņ┤ł ĒÖĢņ¦äļÉśĻ│Ā ņØ┤Ēøä 2015ļģäĻ╣īņ¦Ć ņ¦ĆņåŹņĀüņ£╝ļĪ£ ļ│æ ļ░£ņāØņØ┤ ĒÖĢņ¦äļÉ£ Ļ▓ĮĻĖ░ļÅä Ēżņ▓£ņŗ£ ņåīņ×¼ ņé¼Ļ│╝ļéśļ¼┤ Ļ│╝ņłśņøÉņŚÉņä£ ņØ┤ļ│æņ▓┤ļź╝ ņłśņ¦æĒĢśņśĆļŗż. ņłśņ¦æĻĖ░Ļ░äņØĆ 2014ļģäĻ│╝ 2015ļģä ņ┤Ø 2ļģäņŚÉ Ļ▒Ėņ│É ņŗżņŗ£ĒĢśņśĆĻ│Ā Ļ░£ĒÖöņ¦üņĀä ļŗ©Ļ│äņØĖ 4ņøöļČĆĒä░ ņłśĒÖĢĻĖ░ņØĖ 10ņøöĻ╣īņ¦ĆņśĆņ£╝ļ®░, 2014ļģäņØĆ 7ĒÜī(4ņøö 29ņØ╝, 6ņøö 5ņØ╝, 6ņøö 19ņØ╝, 7ņøö 1ņØ╝, 8ņøö 5ņØ╝, 8ņøö 21ņØ╝ ļ░Å 10ņøö 2ņØ╝), 2015ļģäņØĆ 13ĒÜī(4ņøö 17ņØ╝, 5ņøö 1ņØ╝, 5ņøö 15ņØ╝, 5ņøö 29ņØ╝, 6ņøö 12ņØ╝, 6ņøö 26ņØ╝, 7ņøö 10ņØ╝, 7ņøö 27ņØ╝, 8ņøö 7ņØ╝, 8ņøö 21ņØ╝, 9ņøö 4ņØ╝, 9ņøö 18ņØ╝ ļ░Å 10ņøö 2ņØ╝) ņłśņ¦æĒĢśņśĆļŗż. ņłśņ¦æ ņŗ£ 4ņøöĻ│╝ 5ņøöņØĆ ņé¼Ļ│╝ļéśļ¼┤ņŚÉņä£ ņ¢┤ļ¢ĀĒĢ£ ņ£Āņé¼ ņ”ØņāüļÅä Ļ┤Ćņ░░ĒĢĀ ņłśĻ░Ć ņŚåņ¢┤ Ļ▒┤ņĀä ņżäĻĖ░ ļ░Å ņ×ÄņØä ņ▒äņĘ©ĒĢśņśĆņ£╝ļ®░, ņØ┤ĒøäņŚÉļŖö ņĀäĒśĢņĀüņØĖ ļ¦łļ”äņ”ØņāüņØś ņØ┤ļ│æņĪ░ņ¦üņØ┤ Ļ┤Ćņ░░ļÉśņ¢┤ ņØ┤ļź╝ ņżæņŗ¼ņ£╝ļĪ£ ņłśņ¦æĒĢśņśĆĻ│Ā, 9ņøö ņØ┤ĒøäļŖö ņłśĒÖĢ ļ░Å ņČ£ĒĢśņŗ£ĻĖ░ļĪ£ ņä▒ņłÖĻ│╝ņŗżļ¦īņØä ļīĆņāüņ£╝ļĪ£ ĒĢśņśĆļŗż.
ļ│æņøÉņäĖĻĘĀņØś ļČäļ”¼ļŖö ņłśņ¦æļÉ£ ņŗ£ļŻīļź╝ ice boxņŚÉ ļ│┤Ļ┤ĆĒĢśņŚ¼ ņŗżĒŚśņŗżļĪ£ ņØ┤ļÅÖĒĢ£ Ēøä, 24ņŗ£Ļ░ä ņØ┤ļé┤ņŚÉ ņłśĒ¢ēĒĢśņśĆļŗż. Ļ░ü ņłśņ¦æļÉ£ ĻĮā, Ļ░Ćņ¦Ć, ņ×Ä, ĻĘĖļ”¼Ļ│Ā Ļ│╝ņŗżņØĆ ļ®ĖĻĘĀļÉ£ ļ®┤ļÅäņ╣╝ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņ¦üĻ▓Į 3 mmļĪ£ ņלļØ╝ 70% ņŚÉĒāäņś¼ņŚÉņä£ 5ļČä, ļ®ĖĻĘĀņłś Ļ░ü 5ļČä 3ĒÜī ņäĖņ▓ÖĒĢśņŚ¼ Ļ▒┤ņĪ░ Ēøä, Ļ░ü ņĪ░ņ¦üņØä ļ®ĖĻĘĀņłśņŚÉņä£ ļ¦łņćäĒĢ£ Ēøä, 10ļ░░ ĒؼņäØļ▓Ģņ£╝ļĪ£ 10-3Ļ╣īņ¦Ć ĒؼņäØĒĢśņŚ¼ mannitol glutamate yeast extract (MGY; D-mannitol 10 g, L-glutamic acid 2 g, KH2PO4 0.5 g, NaCl 0.2 g, MgSO4 ┬Ę 7H2O 0.2 g, yeast extract 1 g, agar 15 g/l, pH 7.0) ļ░░ņ¦ĆņŚÉ ļÅäļ¦ÉĒĢśņŚ¼ 28┬░CņŚÉņä£ 24ņŗ£Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ņØ┤ļĢī ņä▒ņłÖĻ│╝ņŗżņØĆ Ļ│╝ņŗżņ×ÉļŻ©ņÖĆ Ļ│╝ņŗżĒæ£ļ®┤ ļ░Å ļé┤ļČĆļĪ£ ļéśļłäņ¢┤ ļČäļ”¼ĒĢśņśĆļŗż. ļ░░ņ¢æ 24ņŗ£Ļ░ä Ēøä ĒśĢņä▒ļÉśļŖö ļ¬©ļōĀ ĻĘĀņ┤ØļōżņØĆ Ēü¼ĻĖ░, ņāē ļ░Å ļ¬©ņ¢æ ļō▒ņØś ņä▒ņāüņŚÉ ļö░ļØ╝ ļÅÖņØ╝ ļ░░ņ¦ĆņŚÉ ņł£ņłśļČäļ”¼ĒĢśņŚ¼ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØś ņä▒ņāüĻ│╝ ļ╣äĻĄÉĒĢśņśĆļŗż.
Colony-PCRņØä ņ£äĒĢ£ ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ė ņäĀļ░£
ņłśņ¦æļÉ£ ņŗ£ļŻīļĪ£ļČĆĒä░ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØś ņŗĀņåŹĒĢ£ Ļ▓ĆņČ£ņØä ņ£äĒĢśņŚ¼, genomic DNA ņČöņČ£Ļ│╝ņĀĢ ņŚåņØ┤ Ļ░ü ĻĘĀņ┤ØļōżļĪ£ļČĆĒä░ ņ¦üņĀæ PCR ļ░śņØæņØä ĒåĄĒĢ£ Ļ▓ĆņČ£ļĪ£ ņłśĒ¢ēļÉśņŚłļŗż. ņØ┤ņŚÉ ņä▒ņāüņØ┤ ļŗżļźĖ ļŗ©ņØ╝ ĻĘĀņ┤Øļōż ņżæņŚÉņä£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀĻ│╝ Ļ░Ćņן ņ£Āņé¼ĒĢ£ ņä▒ņāüņØä ļéśĒāĆļé┤ļŖö ĻĘĀņ┤ØļōżņØä ņØ┤ņÜ®ĒĢśņŚ¼ colony-PCRņÜ® ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņäĀļ░£ĒĢśņśĆļŗż. Colony-PCRņÜ® ĒöäļØ╝ņØ┤ļ©Ė Ēøäļ│┤ĻĄ░ļōżņØĆ ĻĖ░ņĪ┤ņŚÉ genomic DNAļź╝ ņØ┤ņÜ®ĒĢ£ PCR ļ░śņØæņŚÉņä£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©ĖļĪ£ ļ│┤Ļ│ĀļÉ£ Ļ▓āĻ│╝ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļ│æņøÉņä▒ ļ░Å housekeeping ņ£ĀņĀäņ×Éļź╝ ĒÖ£ņÜ®ĒĢśņŚ¼ Ļ░£ļ░£ļÉ£ ĒöäļØ╝ņØ┤ļ©ĖļōżņØä ļīĆņāüņ£╝ļĪ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ Ļ▓ĆņČ£ ĒŖ╣ņØ┤ colony-PCRņÜ® ĒöäļØ╝ņØ┤ļ©ĖļōżņØä ņäĀļ░£ĒĢśņśĆļŗż(Table 1) (Bereswill ļō▒, 1995; Kim ļō▒, 2001a, 2001b; Park ļō▒, 2010; Shrestha ļō▒, 2005b, 2007). ļśÉĒĢ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØś ĻĘĀņ┤Ø ĒŖ╣ņä▒ņØĆ ĒÖöņāüļ│æņøÉņäĖĻĘĀĻ│╝ļÅä ļ¦żņÜ░ ņ£Āņé¼ĒĢśĻĖ░ņŚÉ, ņØ┤ļź╝ ĻĄ¼ļ│äĒĢĀ ņłś ņ׳ļŖö ĒÖöņāüļ│æņøÉĻĘĀ ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©ĖļÅä ĒżĒĢ©ņŗ£ņ╝░ļŗż(Table 1). Colony-PCR ņĪ░Ļ▒┤ņØĆ ļ®ĖĻĘĀņłś 36.5 ╬╝lņŚÉ ļŗ©ņØ╝ ĻĘĀņ┤ØņØä ņØ┤ņæżņŗ£Ļ░£ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņåīļ¤ē ĒśäĒāüĒĢśĻ│Ā 100┬░CņŚÉņä£ 3ļČäĻ░ä Ļ░ĆņŚ┤ĒĢ£ Ēøä, 10├Ś buffer (1 M KCl, 1 M Tris-HCl, pH 8.0, 1 M MgCl2) 5 ╬╝l, 30% DMSO 5 ╬╝l, 20 mM dNTP 0.5 ╬╝l, Ļ░ü 10 ĒéĪ forward ļ░Å reverse ĒöäļØ╝ņØ┤ļ©Ė 1 ╬╝l, Taq polymerase (handmade type) 1 ╬╝lļź╝ ņ▓©Ļ░ĆĒĢśņŚ¼ ņ┤Ø 50 ╬╝l ļČĆĒö╝ņŚÉņä£ ņłśĒ¢ēļÉśņŚłļŗż. ļ░śņØæņŗ£Ļ░äņØĆ 95┬░C 2ļČä Ēøä, 94┬░C 15ņ┤ł, Ļ░ü ĒöäļØ╝ņØ┤ļ©Ė ņóģļźśļ│ä annealing conditions (Table 1), 68┬░C 1ļČä 30ņ┤ł ņĪ░Ļ▒┤ņ£╝ļĪ£ ņ┤Ø 35ĒÜī ļ░śņØæĒĢśņśĆņ£╝ļ®░, ņĄ£ņóģ 68┬░C 10ļČäņ£╝ļĪ£ extensionņØä ņÖäņä▒ĒĢśņśĆļŗż. Coloy-PCR ņé░ļ¼╝ņØś ĒÖĢņØĖņØĆ 1% agarose gel ņĀäĻĖ░ņśüļÅÖņØä ņłśĒ¢ēĒĢśņŚ¼ ethidium bromide (1 mg/ml)ņŚÉ ņŚ╝ņāēņŗ£Ēé© Ēøä UV transilluminatorņŚÉņä£ Ļ░ü ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ė ņóģļźśļ│ä ņśłņāü Ēü¼ĻĖ░ņØś ļ░┤ļō£ņāØņä▒ ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. Ļ░ü ĒöäļØ╝ņØ┤ļ©Ė ņóģļźśļ│ä ĒŖ╣ņØ┤ ļ░┤ļō£ Ļ▓ĆņČ£ ņāØņä▒ ņŚ¼ļČĆļŖö 1995ļģä ņČśņ▓£ņŚÉņä£ ļČäļ”¼ļÉ£ WT3 ĻĘĀņŻ╝(Shrestha ļō▒, 2003) ļ░Å 1996ļģä ļČäļ”¼ĻĘĀņŻ╝ņØĖ Ep1 (Kim ļō▒, 1999)ņØä ļīĆņĪ░ĻĘĀņŻ╝ļĪ£ ĒÖ£ņÜ®ĒĢśņśĆņ£╝ļ®░, ĒÖöņāüļ│æņøÉĻĘĀņŻ╝ļŖö type strainņØĖ ATCC15580ņØä ņé¼ņÜ®ĒĢśņŚ¼ ļ╣äĻĄÉ ļČäņäØĒĢśņśĆļŗż.
Table┬Ā1
Primers used in this study
| ŌĆāPrimers | Sequences 5ŌĆÖ-3ŌĆÖ | Expected | Target | Annealing | Source or reference |
|---|---|---|---|---|---|
| size (bp) | species | condition* | |||
| AMSbL | GCTACCAGCAGGGTGAG | 1,600 | Ea | 49┬░C, 2 min | Bereswill et al., 1995 |
| AMSbR | TCATCACGATGGTGTAG | ||||
| P29A | CGGTTTTTAACGCTGGG | 1,000 | Ea | 52┬░C, 2 min | Kim et al., 2001a |
| P29B | GGGCAAATACTCGGATT | ||||
| EP16A | AGATGCGGAAGTGCTTCG | 730 | Ep | 52┬░C, 2 min | Kim et al., 2001a |
| EPIG2c | ACCGTTAAGGTGGAATC | ||||
| EpERF | GCGGTCATAGTGGCAATGAT | 638 | Ep | 58┬░C, 30 sec | Park et al., 2010 |
| EpERR | GCACCTGCGATGCAAAGATG | ||||
| EpSPF | AAAGGCGCAGGGCCTGATGA | 573 | Ep | 55┬░C, 30 sec | Shrestha et al., 2007 |
| EpSPR | GCACCTGCGATGCAAAGATG | ||||
| hrpNEpF1 | GTGGTTTAACGGGGCTGCTG | 1,100 | Ep | 64┬░C, 15 sec | Shrestha et al., 2005b |
| hrpNEpR1 | AGCTTAAGCCGCGCCCAG | ||||
| CPS1 | CGCGGAAGTGGTGAGAA | 1,200 | Ep | 52┬░C, 15 sec | Kim et al., 2001b |
| CPS2c | GAACAGATGTGCCGAGTA | ||||
| recAF | CAATTTGGCATCGATTTCGTT | 565 | Ep | 55┬░C, 30 sec | This study |
| recAR | AGGGGCAGTAGATGTCATCA | ||||
| dspEF | ATGCCAGCTTTTGCAGGCTGT | 449 | Ep | 55┬░C, 30 sec | This study |
| dspER | TTTCACCTCTGGCGAGTTTG | ||||
| gyrAF | GCTGCAGACCTCTTTTGGTA | 1,099 | Ep | 55┬░C, 30 sec | This study |
| gyrAR | GCTACCGCTAGTCAGCGAAA |
ļ░£ņāØņ¦Ć ņś©ļÅä ņĖĪņĀĢ
Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ļ░£ņāØĻ│╝ ņś©ļÅäņÖĆņØś Ļ┤ĆĻ│äļź╝ ļČäņäØĒĢśĻĖ░ ņ£äĒĢ┤ ņŗ£ļŻīņłśņ¦æ ņןņåīņŚÉ Humidity/Temp. data recorder (MHT-381SD; Lutron Electronic Enterprise Co., Taipei, Taiwan)ļź╝ ņäżņ╣śĒĢśņŚ¼ 10ļČä Ļ░äĻ▓®ņ£╝ļĪ£ ņś©ļÅäļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ņØ╝ņØ╝ ĒÅēĻĘĀņś©ļÅä=[(ņØ╝ ņĄ£Ļ│ĀĻĖ░ņś©+ņØ╝ ņĄ£ņĀĆĻĖ░ņś©)/2]ļĪ£ ņé░ņĀĢĒĢśņŚ¼ ņØ╝ ĒÅēĻĘĀņś©ļÅäņŚÉ ļö░ļźĖ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØņāüĒÖ®ņØä ļČäņäØĒĢśņśĆļŗż.
ņØ┤ļ│æņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ Ļ▒░ļ”¼ņŚÉ ļö░ļźĖ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀ Ļ▓ĆņČ£
2014-2015ļģä ļÅÖņĢł ņØ┤ļ│æņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉ£ ĻĖ░Ļ░ä ļé┤ņØś ņŗ£ļŻīļōż ņżæņŚÉņä£, 2014ļģä 6ņøö 5ņØ╝, 6ņøö 19ņØ╝ ļ░Å 7ņøö 1ņØ╝, ĻĘĖļ”¼Ļ│Ā 2015ļģä 6ņøö 12ņØ╝ ņłśņ¦æļÉ£ ņØ┤ļ│æņżäĻĖ░ļŖö ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦üņØä ļ¬ģĒÖĢĒ׳ ĻĄ¼ļČäĒĢĀ ņłś ņ׳ņŚłļŗż. ņØ┤ņŚÉ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ ņØ┤ļ│æņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ Ļ▒░ļ”¼ņŚÉ ļö░ļØ╝ ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ņ£Āļ¼┤ļź╝ ĒÖĢņØĖĒĢśņśĆļŗż. ņØ┤ ņŗ£ĻĖ░ņŚÉ ņłśņ¦æļÉ£ ņżäĻĖ░ļŖö ļ¬©ļæÉ 2ļģäņāØ ņØ┤ņāü ņżäĻĖ░ļĪ£ ņ¦üĻ▓ĮņØ┤ 1 cm ņØ┤ņāüņØ┤ņ¢┤ņä£ ļ│æņøÉĻĘĀ ļČäļ”¼ļź╝ ņ£äĒĢ£ ļ¦łņćäĻ░Ć ņÜ®ņØ┤ĒĢśņ¦Ć ņĢŖņĢä, ļ®ĖĻĘĀļÉ£ ļ®┤ļÅäņ╣╝ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņלĻ▓ī ļČäņćä Ēøä ļ®ĖĻĘĀņłśņŚÉ ņ╣©ņ¦ĆĒĢśņŚ¼ 10ļČäĻ░ä ņĀĢņ╣śĒĢśĻ│Ā vortaxingĒĢ£ Ēøä, ļŗżļźĖ ņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ņØś ļČäļ”¼Ļ│╝ņĀĢĻ│╝ ļÅÖņØ╝ĒĢśĻ▓ī ļ░░ņ¢æĒĢśņŚ¼ ņāØņןĒĢśļŖö ļŗ©ņØ╝ ĻĘĀņ┤ØņØä ņł£ņłśļČäļ”¼ĒĢśņśĆļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ĒŖ╣ņØ┤ colony-PCR ĒöäļØ╝ņØ┤ļ©Ė
ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņØś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØś Ļ▓ĆņČ£ņØĆ ņł£ņłś ņäĖĻĘĀ DNA ņČöņČ£Ļ│╝ņĀĢ ņŚåņØ┤ ņłśĒ¢ēļÉĀ Ļ▓ĮņÜ░, ņŗ£Ļ░äņĀłņĢĮĻ│╝ ļ╣äņÜ®ņĀłĻ░É ļ®┤ņŚÉņä£ Ēü░ ņØ┤ņØĄņØä ņ¢╗ņØä ņłś ņ׳ļŖö ņĀłņ░©ņØ┤ļŗż. ĻĘĖļ¤¼ļéś ļ░śļō£ņŗ£ ļÅÖļ░śļÉśņ¢┤ņĢ╝ ĒĢĀ ņĪ░Ļ▒┤ņØĆ ņł£ņłś DNAļź╝ ĒÖ£ņÜ®ĒĢśļŖö Ļ▓āņØ┤ ņĢäļŗłļ»ĆļĪ£ colony-PCRņŚÉņä£ļÅä ņØ┤ļōż ĒöäļØ╝ņØ┤ļ©ĖņØś ĒŖ╣ņØ┤ņĀü Ļ▓ĆņČ£ļŖźļĀźņØ┤ ĒÖĢļ│┤ļÉśņ¢┤ņĢ╝ļ¦ī ĒĢ£ļŗż. ņØ┤ņŚÉ ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ĒÖ£ņÜ®ļÉ£ ĒöäļØ╝ņØ┤ļ©ĖļōżņØĆ ĻĖ░ņĪ┤ ļ│┤Ļ│ĀņŚÉņä£ ļ¬©ļæÉ ņČöņČ£ļÉ£ genomic DNAļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ĒŖ╣ņØ┤ņä▒ņØ┤ Ļ▓Ćņ”ØļÉ£ Ļ▓āļōżļĪ£, ņäĖĒżļ¦īņØä Ļ░Ćņ¦ĆĻ│Ā ņłśĒ¢ēĒĢśļŖö colony-PCRņŚÉņä£ņØś ĒÖ£ņÜ®ņä▒ ņŚ¼ļČĆļź╝ Ļ▓Ćņ”ØĒĢ┤ņĢ╝ļ¦ī ĒĢĀ ĒĢäņÜöĻ░Ć ņ׳ņŚłļŗż. 2ņóģļźś(WT3 ļ░Å Ep1)ņØś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņŚÉņä£ļŖö ĒŖ╣ņØ┤ ļ░┤ļō£ļź╝ ņāØņä▒ĒĢ£ ļ░śļ®┤, ĒÖöņāüļ│æņøÉņäĖĻĘĀņŚÉņä£ļŖö ļéśĒāĆļéśņ¦Ć ņĢŖļŖö ĒöäļØ╝ņØ┤ļ©ĖļōżļĪ£ļŖö CPS1/CPS2c, EP16A/EPIG2c, EpERF/EpERR, EpSPF/EpSPR, recAF/recAR, dspEF/dspER, ĻĘĖļ”¼Ļ│Ā gyrAF/gyrARļĪ£ ļéśĒāĆļé¼ļŗż(data not shown). recAF/recAR, dspEF/dspER ļ░Å gyrAF/gyrAR ĒöäļØ╝ņØ┤ļ©ĖļōżņØĆ ļ╣äĒŖ╣ņØ┤ ļīĆņāü ĻĘĀņŻ╝ņØĖ ATCC15580ņŚÉņä£ļÅä ņśłņāü Ēü¼ĻĖ░ņØś ļ░┤ļō£ļŖö ņĢäļŗłņ¦Ćļ¦ī ļŗżļźĖ Ēü¼ĻĖ░ņØś ļ░┤ļō£ļź╝ ņāØņä▒ĒĢśņŚ¼ ņĀ£ņÖĖņŗ£ņ╝░ņ£╝ļ®░, EP16A/EPIG2c ĒöäļØ╝ņØ┤ļ©ĖļŖö WT3ņÖĆ Ep1 ĻĘĀņŻ╝ņŚÉņä£ņØś ĒŖ╣ņØ┤ ļ░┤ļō£ ņāØņä▒ Ļ░ĢļÅäņØś ņ░©ņØ┤Ļ░Ć ņ׳ņŚłĻ│Ā, CPS1/CPS2cļŖö ĒŖ╣ņØ┤ ļ░┤ļō£ņØś Ēü¼ĻĖ░Ļ░Ć 1.2 kbļĪ£ ļŗżņåī Ēü░ ļ¼ĖņĀ£ņĀÉņØ┤ ņ׳ņŚłļŗż(data not shown). Ļ░Ćņן ĒÜ©ņ£©ņĀüņØĖ colony-PCRņÜ® Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©ĖļŖö EpERF/EpERRĻ│╝ EpSPF/EpSPRņØś 2ņóģļźśņśĆļŗż(Fig. 1). ņØ┤ļōżņØĆ ļ¬©ļæÉ enterobacterial repetitive intergenic consensus (ERIC) ļČĆņ£ä ņ£Āļל ĒŖ╣ņØ┤ ņŚ╝ĻĖ░ņä£ņŚ┤ ņ¦ĆņŚŁņŚÉņä£ ņĀ£ņ×æļÉ£ ĒöäļØ╝ņØ┤ļ©ĖļōżļĪ£(Park ļō▒, 2010; Shrestha ļō▒, 2007), colony-PCRņŚÉņä£ļÅä ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ĒŖ╣ņØ┤ņä▒ņØä ļéśĒāĆļāäņØä ņĢī ņłś ņ׳ņŚłļŗż. ņØ┤ņŚÉ ņĄ£ņóģņĀüņ£╝ļĪ£ ņØ┤Ēøä ņŗżĒŚśņŚÉņä£ļŖö EpSPF/EpSPR ĒöäļØ╝ņØ┤ļ©Ė ņīŹņØä ņØ┤ņÜ®ĒĢśņŚ¼ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀ Ļ▓ĆņČ£ņØä ņłśĒ¢ēĒĢśņśĆļŗż.
Fig.┬Ā1
Confirmation of expected size by colony-PCR assay with EpERF/EpERR (A) and EpSPF/EpSPR (B) primer sets. WT3 and Ep1, and ATCC15580 are representative strains for Erwinia pyrifoliae and Erwinia amylovora, respectively, tested in this study. M, marker of 1 kb ladder DNA. Expected band sizes are shown at the left.

ņāØņ£ĪĻĖ░Ļ░ä ņś©ļÅäņŚÉ ļö░ļźĖ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØņāüĒÖ®
2013ļģä ņØ┤Ēøä ņĪ░ņé¼ĻĖ░Ļ░ä(2014-2015ļģä) ļÅÖņĢł 4ņøöļČĆĒä░ 10ņøöĻ╣īņ¦Ć ņłśņ¦æļÉ£ ļ¬©ļōĀ ņŗ£ļŻīļĪ£ļČĆĒä░ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØä ļČäļ”¼ĒĢśĻ│Ā colony-PCR ļ░®ļ▓Ģņ£╝ļĪ£ ļ│æņøÉņäĖĻĘĀņØä Ļ▓ĆņČ£ĒĢśņśĆļŗż. 2014ļģä ņŗ£ļŻīņŚÉņä£ļŖö 6-7ņøö ņŗ£ļŻī(6ņøö 5ņØ╝, 6ņøö 19ņØ╝, 7ņøö 1ņØ╝)ņŚÉņä£ļ¦ī Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņŚłļŖöļŹ░, ņØ┤ļŖö ņØ╝ ĒÅēĻĘĀņś©ļÅä 20┬░C-25┬░C ļ▓öņ£äņśĆļŗż. 2015ļģäņØĆ 5ņøö ļ¦É(5ņøö 29ņØ╝)ļČĆĒä░ 6ņøö ņżæņł£(6ņøö 12ņØ╝)ņŚÉļ¦ī ļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░, ņś©ļÅäļ▓öņ£äļŖö 25┬░C-30┬░CņśĆļŗż. ĒĢ£ĒÄĖ 7ņøö ņżæņł£ ņØ┤Ēøä ļ│æņ¦ĢņØ┤ Ļ┤Ćņ░░ļÉ£ ņżäĻĖ░ņÖĆ ņ×ÄņØ┤ ņĀ£Ļ▒░ļÉśņ¦Ć ņĢŖĻ│Ā ĻĘĖļīĆļĪ£ ļ│┤ņĪ┤ļÉśņ¢┤ ņ׳ļŖö ņØ┤ļ│æ ņé¼Ļ│╝ļéśļ¼┤ņŚÉņä£ ņØ┤ļ│æņ▓┤ļź╝ ņłśņ¦æĒĢśņŚ¼ Ļ▓ĆņČ£ĒĢ£ Ļ▓░Ļ│╝, 2014ļģäĻ│╝ 2015ļģä ļ¬©ļæÉ ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ļŖö ļČäļ”¼ ļ░Å colony-PCR ļ¬©ļæÉņŚÉņä£ ĒÖĢņØĖĒĢĀ ņłś ņŚåņŚłļŗż. ļö░ļØ╝ņä£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ņĄ£ņĀü ļ░£ņāØ ĒÅēĻĘĀņś©ļÅäļŖö 25┬░C ļé┤ņÖĖļĪ£ ņāØĻ░üļÉśņ¢┤ Shrestha ļō▒(2005a)ņØ┤ ļ│┤Ļ│ĀĒĢ£ ļé┤ņÜ®Ļ│╝ ņ£Āņé¼ĒĢśņśĆņ£╝ļ®░, ņØ┤Ēøä ņØ╝ ĒÅēĻĘĀņś©ļÅäĻ░Ć 30┬░C ņØ┤ņāü ņ£Āņ¦ĆļÉĀ Ļ▓ĮņÜ░ņŚÉļŖö Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØĆ ņś©ļÅäņĀüņØæ ļŖźļĀźņØ┤ Ļ░ÉņåīĒĢśĻ│Ā, ņØ┤ļŖö ļ░ĆļÅäĻ░ÉņåīņØś ņøÉņØĖņØ┤ ļÉśļŖö Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉśņŚłļŗż. ņØ┤ļ¤¼ĒĢ£ Ļ│Āņś©ņĪ░Ļ▒┤ņŚÉņä£ņØś ļ│æņøÉņäĖĻĘĀņØś ņāØņĪ┤ņĀäļץ ļ░Å ņøöļÅÖĻ│╝ ĒĢśļÅÖņØä ņ£äĒĢ£ ņØ┤ļÅÖņĀäļץ ĻĘĖļ”¼Ļ│Ā ņןņåīņŚÉ ļīĆĒĢ┤ņä£ļŖö Ē¢źĒøä ļŹö ņŗ¼ļÅä Ļ╣ŖņØĆ ņŚ░ĻĄ¼Ļ░Ć ņØ┤ļŻ©ņ¢┤ņĀĖņĢ╝ļ¦ī ĒĢ£ļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż.
ĒÜ©ņ£©ņĀü Ļ░Ćņ¦Ćņ╣śĻĖ░ļź╝ ņ£äĒĢ£ ņØ┤ļ│æņ▓┤ ņĀ£Ļ▒░
ļ│æņ¦ĢņØä ņ£ĪņĢłņ£╝ļĪ£ Ļ┤Ćņ░░ĒĢĀ ņłś ņ׳ļŖö ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ░äņØś ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ņ£Āļ¼┤ļź╝ Ļ┤Ćņ░░ĒĢ£ Ļ▓░Ļ│╝, 2014ļģä 6ņøö 15ņØ╝ ņŗ£ļŻīņŚÉņä£ļŖö ļ¬©ļōĀ ņØ┤ļ│æņĪ░ņ¦ü ļé┤ņØś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀĻ│╝ ņ£Āņé¼ĒĢ£ ņä▒ņāüņØä ļéśĒāĆļé┤ļŖö ĻĘĀņ┤ØļōżņØĆ 573 bpņØś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀ ĒŖ╣ņØ┤ ļ░┤ļō£ļź╝ ņāØņä▒ĒĢśņśĆļŗż. ļśÉĒĢ£ ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 10 cm ļ░Å 20 cm Ļ▒░ļ”¼ņØś Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ ļČäļ”¼ļÉ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ņ£Āņé¼ ĻĘĀņ┤Øļōż ļ¬©ļæÉ ĒŖ╣ņØ┤ ļ░┤ļō£ļź╝ ļéśĒāĆļé┤ņ¢┤, ņØ┤ļ│æņĪ░ņ¦üņ£╝ļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ 20 cmĻ╣īņ¦Ć Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ ņĪ┤ņ×¼ĒĢĀ ņłś ņ׳ņØīņØ┤ ņ”Øļ¬ģļÉśņŚłļŗż(Fig. 2A). ĻĘĖļ¤¼ļéś 20 cm ņØ┤ņāüņØś Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ņØś Ļ▓ĆņČ£ņØĆ ņØ┤ļŻ©ņ¢┤ņ¦Ćņ¦Ć ņĢŖņĢä, 6ņøö 19ņØ╝Ļ│╝ 7ņøö 1ņØ╝ ņŗ£ļŻīļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 20 cm ņØ┤ņāüņØś Ļ▒░ļ”¼ņŚÉņä£ļÅä ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ņ£Āļ¼┤ļź╝ Ļ▓ĆņĀĢĒĢśņśĆļŗż. ļ©╝ņĀĆ 6ņøö 19ņØ╝ ņŗ£ļŻīņŚÉņä£ļŖö Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ ņĄ£ņן 81 cmĻ╣īņ¦Ć Ļ▓ĆņĀĢĒĢśņśĆļŖöļŹ░, ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 13 cm Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ļŖö 6ņøö 5ņØ╝ ņŗ£ļŻīņÖĆ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ ļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░, 20 cm ņØ┤ņāüņØś Ļ░äĻ▓®ņØĖ 26 cm, 55 cm ļ░Å 81 cm Ļ░äĻ▓®ņØś ņĪ░ņ¦üņŚÉņä£ļŖö Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśļŗż(Fig. 2B). ļśÉĒĢ£ 7ņøö 1ņØ╝ ņŗ£ļŻīņŚÉņä£ļÅä ņØ┤ļ│æņĪ░ņ¦üņŚÉņä£ļŖö ļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉ£ ļ░śļ®┤, Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 26 cm Ļ░äĻ▓®ņØś Ļ▒┤ņĀäņĪ░ņ¦üņŚÉņä£ļŖö Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢä ņ£ĀņØśņä▒ ņ׳ļŖö Ļ▓░Ļ│╝ļź╝ ļ│┤ņŚ¼ ņŻ╝ņŚłļŗż(Fig. 2C). 2015ļģä ņŗ£ļŻīņŚÉņä£ļÅä ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 20 cm ņØ┤ļé┤ņØś Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ļŖö ļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░, ņØ┤ņāüņØś Ļ░äĻ▓®ņØĖ 30 cm Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ļŖö Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØ┤ Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśļŗż(Fig. 2D). 2014ļģä 6ņøö 5ņØ╝ ņŗ£ļŻīņŚÉņä£ Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ 10 cm Ļ░äĻ▓®ņØś Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņŚÉņä£ ļČäļ”¼ļÉ£ ĻĘĀņ┤Ø(Fig. 2A)Ļ│╝ 2015ļģä 6ņøö 12ņØ╝ ņŗ£ļŻīņØś ņØ┤ļ│æņĪ░ņ¦ü ļé┤ņŚÉņä£ ļČäļ”¼ļÉ£ ĻĘĀņ┤Ø(Fig. 2D)ņŚÉņä£ļŖö colony-PCR Ļ▓ĆņČ£ļ░®ļ▓ĢņŚÉ ņØśĒĢ┤ņä£ ĒŖ╣ņØ┤ ļ░┤ļō£ļź╝ ĒśĢņä▒ĒĢśņ¦Ć ņĢŖņĢśļŖöļŹ░, ņØ┤ļŖö ņé¼Ļ│╝ļéśļ¼┤ ņ×ÄĻ│╝ ņżäĻĖ░ ļé┤ņÖĖļČĆņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļČĆņāØņä▒ ņäĖĻĘĀ ņżæņŚÉņä£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀĻ│╝ ņ£Āņé¼ĒĢ£ ņä▒ņāüņØä ļéśĒāĆļé┤ļŖö ĻĘĀņ┤ØņØ┤ ļŗżņłś ļČäļ”¼ļÉśņŚłĻĖ░ ļĢīļ¼ĖņØ┤ļŗż. ĒĢ£ĒÄĖ ņłśĒÖĢĻĖ░ņŚÉ ņä▒ņłÖĻ│╝ļĪ£ļČĆĒä░ ļČäļ”¼ļÉ£ ņäĖĻĘĀļōżņØĆ ļ│æņøÉņäĖĻĘĀĻ│╝ ņä▒ņāüņØ┤ ļ¦żņÜ░ ļŗżļź┤Ļ│Ā, colony-PCR Ļ▓ĆņČ£ļ░®ļ▓ĢņŚÉ ņØśĒĢ┤ņä£ļÅä ĒŖ╣ņØ┤ ļ░┤ļō£Ļ░Ć Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢä, Ļ░ÉņŚ╝ ņé¼Ļ│╝ļéśļ¼┤ņŚÉņä£ ņłśĒÖĢĒĢ£ ņä▒ņłÖ ņé¼Ļ│╝ņŚÉņä£ļŖö ļ│æņøÉņäĖĻĘĀņØ┤ ņĪ┤ņ×¼ĒĢśņ¦Ć ņĢŖļŖö Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż(data not shown).
Fig.┬Ā2
(A: left panel) The diagram of collected sample at June 5th, 2014, to determine distance of existing Erwinia pyrifoliae from infected region on twig. I-1: 60 cm infected distance from border line between symptomatic and asymptomatic regions as specialized form of shoot blight and canker formation with crack. I-2: 30 cm infected distance from border line. I-3: nearby region of symptom from margin. H-1: nearby asymptomatic region from margin. H-2: 10 cm healthy distance from border line. H-3: 20 cm healthy distance from border line. (A: right panel) Detection of E. pyrifoliae by colony-PCR with EpSPF/EpSPR primer pair. Lanes 1-3, colonies isolated from I-1; lanes 4 and 5, colonies isolated from I-2; lanes 6-8, colonies isolated from I-3; lanes 9-13, colonies isolated from H-1; lanes 14 and 15, colonies isolated from H-2; lanes 16-19, colonies isolated from H-3; lanes 20-22, colonies from infected fruit. (B: left panel) Sites for determining E. pyrifoliae from collected sample at June 19th, 2014. I-1: Infected region. H-1: 13 cm healthy distance from border line. H-2: 26 cm healthy distance from border line. H-3: 55 cm healthy distance from border line. H-4: 81 cm healthy distance from border line. (B: right panel) Lanes 1-4, colonies isolated from I-1; lane 5, colony isolated from H-1; lanes 6 and 7, colonies isolated from H-2; lanes 8-10, colonies isolated from H-3; lanes 11-13, colonies isolated from H-4. (C: left panel) Sites for determining E. pyrifoliae from collected sample at July 1st, 2014. I-1: 60 cm infected distance from border line. I-2: 40 cm infected distance from border line. I-3: 20 cm infected distance from border line. I-4: nearby region of symptom from margin. H-1: 26 cm healthy distance from border line. (C: right panel) Lanes 1 and 2, colonies isolated from I-1; lane 3, colony isolated from I-2; lane 4, colony isolated from I-3; lane 5, colony isolated from I-4; lanes 6-9, colonies isolated from H-1. (D: left panel) Sites for determining E. pyrifoliae from collected sample at June 12th, 2015. I-1: 20 cm infected distance from border line. I-2 and I-3: nearby region of symptom from margin. H-1: 10 cm healthy distance from border line. H-2: 20 cm healthy distance from border line. H-3: 30 cm healthy distance from border line. (D: right panel) Lane 1, colony isolated from I-1; lane 2, colony isolated from I-2; lane 3, colony isolated from I-3; lanes 4-6, colonies isolated from H-1; lanes 7 and 8, colonies isolated from H-2; lane 9, colony isolated from H-3. Ep, E. pyrifoliae Ep1; Ea, E. amylovora ATCC15580; water, negative control. M, marker of 1 kb ladder DNA with 500 bp size at the left.
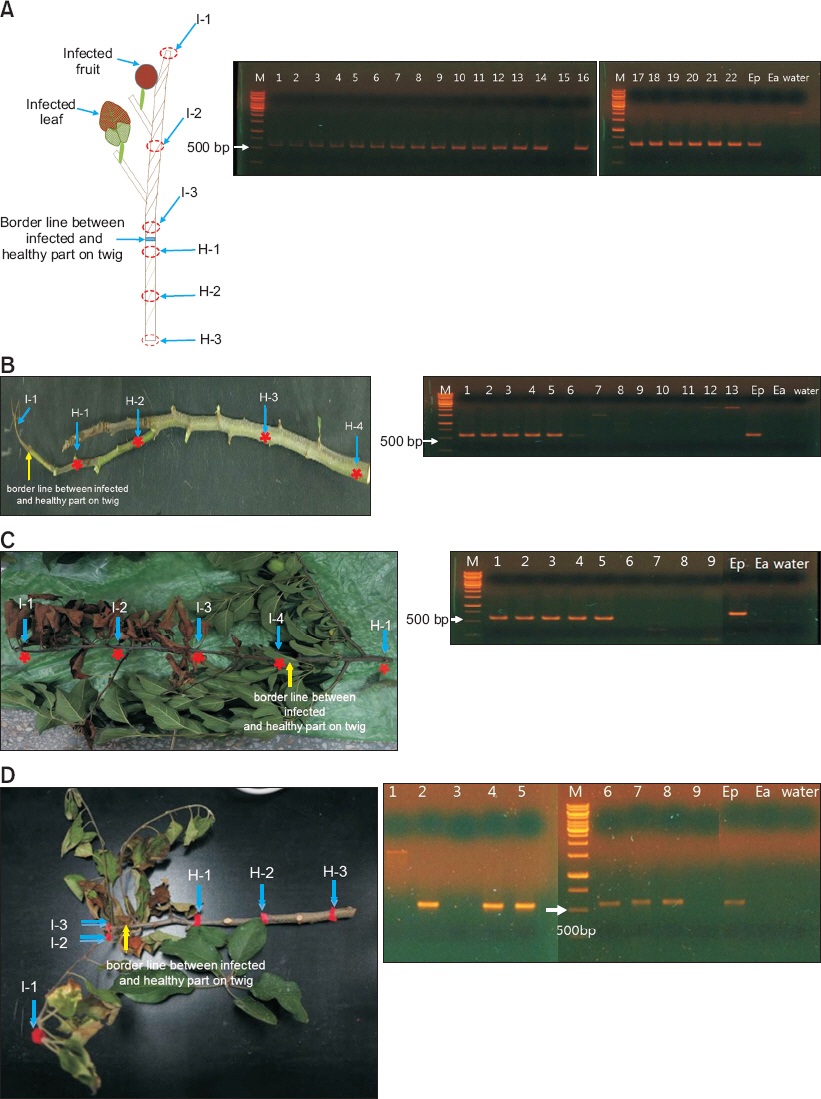
Ļ▓░ļĪĀņĀüņ£╝ļĪ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØĆ ņ£ĪņĢłņ£╝ļĪ£ ļ│æņ¦ĢņØ┤ Ļ┤Ćņ░░ļÉśļŖö ņØ┤ļ│æņĪ░ņ¦ü(ņ×Ä, ņ£ĀĻ│╝ ļ░Å ņżäĻĖ░) ļé┤ņŚÉņä£ Ļ▓ĆņČ£ļÉśņŚłņ£╝ļ®░, ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ 20 cm ņØ┤ņāüņØś Ļ░äĻ▓®ņŚÉņä£ļŖö Ļ▓ĆņČ£ļÉśņ¦Ć ņĢŖņĢśļŗż. ļö░ļØ╝ņä£ ņé¼Ļ│╝ļéśļ¼┤ ņāØņ£Ī ņ┤łņżæĻĖ░ņŚÉ ĒĢ┤ļŗ╣ĒĢśļŖö 5ņøö ļ¦ÉļČĆĒä░ 7ņøö ņ┤łņł£ ņŗ£ĻĖ░ņŚÉ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØ┤ ļ░£ņāØĒĢĀ Ļ▓ĮņÜ░ ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│äļ®┤ņ£╝ļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü 20 cm ņØ┤ņāüņØś Ļ░äĻ▓®ņØä ļæÉĻ│Ā Ļ░Ćņ¦Ćņ╣śĻĖ░ļź╝ ĒĢ£ļŗżļ®┤ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ ļ│æņØś ĒÖĢņé░ņØä ņ¢ĄņĀ£ĒĢĀ ņłś ņ׳ļŗżĻ│Ā ĒīÉļŗ©ļÉśņŚłļŗż. ņØ┤ļŖö Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æĻ│╝ ņ£Āņé¼ĒĢ£ ļ│æņ¦ĢņØä ļéśĒāĆļé┤ļŖö ĒÖöņāüļ│æņØś ĒÖĢņé░ ļ░®ņ¦Ć ļ░Å ļ│æņøÉņäĖĻĘĀņØś ļ░ĆļÅäļź╝ Ļ░Éņåīņŗ£ĒéżĻĖ░ ņ£äĒĢ┤ ņĀäļץņ£╝ļĪ£, ņØ┤ļ│æĻ░Ćņ¦ĆļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ 20-30 cm ņØ┤ņāüņØä ņĀ£Ļ▒░ĒĢĀ Ļ▓ĮņÜ░ ļ¦żņÜ░ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ Ļ┤Ćļ”¼ĒĢĀ ņłś ņ׳ļŗżļŖö ļ│┤Ļ│ĀņÖĆ ņ£Āņé¼ĒĢ£ Ļ▓░Ļ│╝ņØ┤ļŗż(Johnson, 2000; Sundin, 2014; Wilcox, 1994). ļśÉĒĢ£ ĒÖöņāüļ│æņØś Ļ▓ĮņÜ░ 10% ļØĮņŖż ņÜ®ņĢĪ ļśÉļŖö ņŚÉĒāäņś¼ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņĀäņĀĢļÅäĻĄ¼ļź╝ ņåīļÅģĒĢśĻ│Ā, Ļ░Ćņ¦Ćņ╣śĻĖ░ ņŗ£ĻĖ░ļŖö ļ│æņøÉņäĖĻĘĀņØ┤ Ē£┤ļ®┤ĻĖ░ņŚÉ ņ׳ļŖö 2-3ņøöņØ┤ Ļ░Ćņן ņĀüļŗ╣ĒĢśļ®░ ņāØņ£ĪĻĖ░ņŚÉļŖö Ļ░ĢņÜ░ 24ņŗ£Ļ░ä ĒøäĻ░Ć ĒÜ©ņ£©ņĀüņØ┤ļØ╝ļŖö ļ│┤Ļ│ĀņŚÉ ļö░ļØ╝ņä£, Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ņØ┤ļ│æņ▓┤ ņĀ£Ļ▒░ ļśÉĒĢ£ ņ£Āņé¼ĒĢ£ ņŗ£ĻĖ░ņÖĆ ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ ņłśĒ¢ēļÉśļŖö Ļ▓āņØ┤ ļ░öļ×īņ¦üĒĢśļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż.
ļö░ļØ╝ņä£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØś ļ░£ņāØ ņŗ£ ņØ┤ļ│æņ▓┤ļĪ£ļČĆĒä░ Ļ▒┤ņĀäņĪ░ņ¦ü ņżäĻĖ░ļĪ£ 20 cm ņØ┤ņāüņŚÉņä£ Ļ░Ćņ¦Ćņ╣śĻĖ░ņØś Ļ▓ĮņóģņĀü Ļ┤Ćļ”¼ļ░®ļ▓ĢņØĆ, 2015ļģä ļåŹņ┤īņ¦äĒØźņ▓Ł Ļ│╝ņłś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ņśłņ░░ļ░®ņĀ£ņé¼ņŚģ ņ¦Ćņ╣©ņŚÉ ļö░ļØ╝ ņé¼Ļ│╝ļŖö ņØ┤ļ│æņŻ╝Ļ░Ć 10% ņØ┤ņāüņØ╝ Ļ▓ĮņÜ░ ĒÅÉņøÉĒĢśĻ│Ā 10% ņØ┤ĒĢśņØ╝ ļĢīļŖö ņØ┤ļ│æņŻ╝ ļ░Å ņØĖņĀæ 8ņŻ╝ļź╝ ņĀ£Ļ▒░ĒĢ┤ņĢ╝ļ¦ī ĒĢśļŖö ņåÉņŗżņØä ņżäņØ┤ļ®┤ņä£ ļÅÖņŗ£ņŚÉ ĒÜ©ņ£©ņĀüņ£╝ļĪ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØä Ļ┤Ćļ”¼ĒĢĀ ņłś ņ׳ļŖö ļ░®ņĢłņ£╝ļĪ£ ņĀ£ņĢłĒĢśļŖö ļ░öņØ┤ļŗż.
ņÜöņĢĮ
E. pyrifoliaeņŚÉ ņØśĒĢ£ Ļ│╝ņłś Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØĆ ĻĄŁļé┤ņŚÉņä£ 1995ļģä ņĄ£ņ┤ł ļ░£ņāØņØ┤ļל 2016ļģäĻ╣īņ¦Ć ĻŠĖņżĆĒ׳ ļ░£ņāØĒĢśņŚ¼ Ļ│╝ņłśļåŹĻ░ĆņŚÉ Ēö╝ĒĢ┤ļź╝ ņŻ╝Ļ│Ā ņ׳ļŖö ņäĖĻĘĀļ│æĒĢ┤ņØ┤ļŗż. Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØļåŹĻ░ĆņØś ĒÅÉņøÉņĪ░ņ╣ś ļ░Å Ļ│ĄņĀüļ░®ņĀ£ļĪ£ ņØĖĒĢ£ Ļ▓ĮņĀ£ņĀü Ēö╝ĒĢ┤ Ļ░Éņåīļź╝ ņ£äĒĢśņŚ¼, ļ│æņ¦ĢņØ┤ Ļ┤Ćņ░░ļÉśļŖö ņØ┤ļ│æņĪ░ņ¦ü ļ░Å Ļ▒┤ņĀäņĪ░ņ¦ü ļé┤ņØś ļ│æņøÉņäĖĻĘĀņØä Ļ▓ĆņČ£ĒĢśņŚ¼ ĒÜ©ņ£©ņĀü Ļ┤Ćļ”¼ļ░®ņĢłņØä ļ¦łļĀ©ĒĢśĻ│Āņ×É ļ│Ė ņŚ░ĻĄ¼ļź╝ ņłśĒ¢ēĒĢśņśĆļŗż. Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀņØś Ļ▓ĆņČ£ņØĆ genomic DNA ņČöņČ£Ļ│╝ņĀĢņØä ņāØļץĒĢ£ ņł£ņłś ĻĘĀņ┤Øļ¦īņØä ņØ┤ņÜ®ĒĢśļŖö colony-PCRņØä ņØ┤ņÜ®ĒĢśņśĆņ£╝ļ®░, ņØ┤ļź╝ ņ£äĒĢ┤ ERIC ņ¦ĆņŚŁņŚÉņä£ ņĀ£ņ×æļÉ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņøÉņäĖĻĘĀ ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ė EpSPF/EpSPR ĒöäļØ╝ņØ┤ļ©Ė ņīŹņØä ņäĀļ░£ĒĢśņśĆļŗż. ĒŖ╣ņØ┤ ĒöäļØ╝ņØ┤ļ©Ėļź╝ ĒÖ£ņÜ®ĒĢ£ colony-PCR ļ░®ļ▓Ģņ£╝ļĪ£ 2014-2015ļģä 4-10ņøöĻ╣īņ¦Ć ņé¼Ļ│╝ļéśļ¼┤ ņāØņ£ĪĻĖ░Ļ░ä ļÅÖņĢł Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æ ļ░£ņāØņāüĒÖ®ņØä ļ¬©ļŗłĒä░ļ¦üĒĢ£ Ļ▓░Ļ│╝, 25┬░C ņØ╝ ĒÅēĻĘĀņś©ļÅä ĻĖ░Ļ░äņØĖ 5ņøö ņżæņł£ļČĆĒä░ 7ņøö ņ┤łņł£Ļ╣īņ¦Ć ļ░£ļ│æņØ┤ Ļ░Ćņן ļ╣łļ▓łĒĢśņśĆļŗż. ļ░£ļ│æĻ░Ćņ¦Ć ļé┤ ļ│æņøÉņäĖĻĘĀņØś ņĪ┤ņ×¼ņ£Āļ¼┤ Ļ▓ĆņĀĢ Ļ▓░Ļ│╝ ļ│æņ¦Ģ ļČĆņ£äņÖĆ ņØ┤ļĪ£ļČĆĒä░ 20 cm ļé┤ Ļ▒┤ņĀäņĪ░ņ¦üņŚÉņä£ļ¦ī ļ│æņøÉņäĖĻĘĀņØ┤ ņ¦ĆņåŹņĀüņ£╝ļĪ£ Ļ▓ĆņČ£ļÉśņŚłļŗż. ļö░ļØ╝ņä£ ņØ┤ļ»Ė ļ░£ņāØĒĢ£ Ļ░Ćņ¦ĆĻ▓ĆņØĆļ¦łļ”äļ│æņØä ĒÜ©ņ£©ņĀüņ£╝ļĪ£ Ļ┤Ćļ”¼ĒĢśĻĖ░ ņ£äĒĢ┤ ņØ┤ļ│æņĪ░ņ¦üĻ│╝ Ļ▒┤ņĀäņĪ░ņ¦ü Ļ▓ĮĻ│ä ļČĆņ£äļĪ£ļČĆĒä░ 20 cm ņØ┤ņāüņŚÉņä£ Ļ░Ćņ¦Ćņ╣śĻĖ░ļź╝ ĒĢśļŖö Ļ▓āņØ┤ ļ¦żņÜ░ ņĀüņĀłĒĢĀ Ļ▓āņ£╝ļĪ£ ĒīÉļŗ©ļÉ£ļŗż.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






