



ņä£ ļĪĀ
Ļ│żņČ®ņØĆ ņ£ĀĻĖ░ļ¼╝ņØä ņ¦üņĀæ ļČäĒĢ┤ĒĢśĻ▒░ļéś ĒØĪņłśĒĢśļŖö ņåīĒÖöĻĖ░Ļ┤ĆņØ┤ ļ░£ļŗ¼ļÉśņ¢┤ ņ׳ņ¦Ć ņĢŖņĢä ņןļé┤ ļ»ĖņāØļ¼╝ņØś ļÅäņøĆņØä ļ░øņ£╝ļ®░, ņØ┤ļŖö ņןļé┤ņŚÉņä£ ļ╣ĀļźĖ ņ¦äĒÖöņÖĆ ĒĢ©Ļ╗ś ĻĘ╣ĒĢ£ ņä£ņŗØ ĒÖśĻ▓ĮņŚÉņä£ņØś ņĀüņØæņØä Ļ░ĆļŖźĒĢśĻ▓ī ĒĢ£ļŗż. ņØ┤ļ¤¼ĒĢ£ Ļ│żņČ®ņØś ņןļé┤ņŚÉļŖö ļŗżņ¢æĒĢ£ ļ»ĖņāØļ¼╝ņØ┤ ņä£ņŗØĒĢśĻ│Ā ņ׳ņ£╝ļ®░, Ļ│żņČ®Ļ│╝ ņןļé┤ ļ»ĖņāØļ¼╝ ņé¼ņØ┤ņŚÉ ņ£ĀņØĄĒĢ£ ņāüĒśĖņ×æņÜ®ņØä ĒåĄĒĢ┤ Ļ│ĄņāØĻ┤ĆĻ│äļź╝ ņ£Āņ¦ĆĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Rajagopal, 2009). ņØ┤ļĀćļō» Ļ│żņČ® ņןļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö Ļ│ĄņāØņ×ÉņŚÉ ļīĆĒĢ£ ņ×Āņ×¼ņĀüņØĖ ņŚŁĒĢĀņŚÉ Ļ┤ĆĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ĒÖ£ļ░£Ē׳ ņØ┤ļŻ©ņ¢┤ņ¦ĆĻ│Ā ņ׳ļŗż(Genta ļō▒, 2006). ņĄ£ĻĘ╝ ņŚ░ĻĄ¼ņŚÉņä£ Shao ļō▒(2017)ņØĆ ļŗ┤ļ░░Ļ▒░ņäĖļ»Ėļéśļ░®ņØś Ļ│ĄņāØņ×ÉņØĖ ņןļé┤ĻĄ¼ĻĘĀņØĆ ņÖĖļČĆļĪ£ļČĆĒä░ ņ£Āņ×ģļÉśļŖö ņ£ĀĒĢ┤ĻĘĀņØä ņ¢ĄņĀ£ĒĢśļŖö ĒĢŁĻĘĀļ¼╝ņ¦łņØä ļČäļ╣äĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż. Ļ│żņČ® ņןļé┤ ļ»ĖņāØļ¼╝ņØ┤ ligninase, cellulase, lipase, chitinase ļō▒ņØś Ļ│ĀĒÜ©ņ£©ņØś ĒÜ©ņåīļź╝ ņāØņä▒ĒĢ£ļŗżļŖö ļ¦ÄņØĆ ļ│┤Ļ│ĀĻ░Ć ņ׳ņ£╝ļ®░, ņØ┤ļŖö ņé░ņŚģņĀü ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż(Kato ļō▒, 1998; Park ļō▒, 2007; Watanabe ļō▒, 1998). ņØ┤ņ▓śļ¤╝ ņןļé┤ ļ»ĖņāØļ¼╝ņØĆ Ļ│ĀĒÜ©ņ£©ņØś ĒÜ©ņåīļź╝ ņāØņé░ĒĢśĻ│Ā ļīĆņé¼Ļ│╝ņĀĢņŚÉ Ļ┤ĆņŚ¼ĒĢĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ļ®┤ņŚŁņ▓┤Ļ│äņŚÉļÅä Ļ┤ĆņŚ¼ĒĢśļ®░, ņāØņןĻ│╝ ļ░£ņ£ĪņŚÉ ĒĢäņłśņĀüņØĖ ņĢäļ»ĖļģĖņé░ņØ┤ļéś ļ╣äĒāĆļ»╝ ļō▒ņØä ĒĢ®ņä▒ĒĢśļŖö Ļ▓āņ£╝ļĪ£ ļ░ØĒśĆņĪīļŗż(Moon ļō▒, 2011).
Ļ│żņČ®ļ│æņøÉņäĖĻĘĀ Bacillus thuringiensis, Photorhabdus spp., Xenorhabdus spp.ļŖö ļéśļ╣ä ļ¬®, Ēīīļ”¼ ļ¬®, ļö▒ņĀĢļ▓īļĀł ļ¬®, ņäĀņČ® ļō▒ņŚÉ ņé¼ņÜ®ļÉśļŖö ņāØļ¼╝ļåŹņĢĮņ£╝ļĪ£ ņÜ░ņłśĒĢ£ ņé┤ņČ®ĒÖ£ņä▒ņ£╝ļĪ£ ļåŹņŚģĒÖśĻ▓ĮņŚÉ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŗż(LaceyņÖĆ Georgis, 2012; Sanahuja ļō▒, 2011). ņĄ£ĻĘ╝ ņŚ░ĻĄ¼ņŚÉņä£ Ruffner ļō▒(2013)Ļ│╝ Kupferschmied ļō▒(2013)ņØĆ Pseudomonas spp.ņØś ņÜ░ņłśĒĢ£ ņé┤ņČ®ĒÖ£ņä▒ņØä ļ│┤Ļ│ĀĒĢśņśĆļŗż. Pseudomonas spp.ļŖö ņŗØļ¼╝ ĻĘ╝ĻČīņŚÉ ņĀĢņ░®ĒĢśņŚ¼ ļŗżņ¢æĒĢ£ ņŗØļ¼╝ ļ│æņøÉĻĘĀņØä ņ¢ĄņĀ£ĒĢśļ®░ ņ×æļ¼╝ņØś ņāØņ£ĪņØä ņ┤ēņ¦äņŗ£ņ╝£ņä£ ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņØś ņÜ░ņłśņä▒ņŚÉ Ļ┤ĆĒĢ£ ļ¦ÄņØĆ ļ│┤Ļ│ĀĻ░Ć ņ׳ļŗż(De Weger ļō▒, 1986; Kumar ļō▒, 2005).
ņāüņŚģņĀüņ£╝ļĪ£ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŖö ņŗØļ¼╝ļ│æ Ļ┤Ćļ”¼ņÜ® ļ»ĖņāØļ¼╝ ņĀ£ņĀ£ļŖö ņŻ╝ļĪ£ Bacillus spp., Paenibacillus spp., Trichoderma spp., Pseudomonas spp., Streptomyces spp. ļō▒ņØ┤ ņ׳ņ£╝ļ®░, ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØś ĒāÉņāēņØĆ ņŚĮĻČī, ĻĘ╝ĻČī, ĒåĀņ¢æņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ļÉśņ¢┤ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ļŗż(Andrews, 1992; Son ļō▒, 2014; Xu ļō▒, 2016). ņØ┤ļ¤¼ĒĢ£ ļ»ĖņāØļ¼╝ņØĆ ņŗØļ¼╝ņØś ļ┐īļ”¼ļéś ņ×ÄņŚÉ ņĀĢņ░®ĒĢśņŚ¼ ņŗØļ¼╝ļ│æņøÉĻĘĀņØä ņ¢ĄņĀ£ĒĢĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņ×æļ¼╝ņØś ņāØņ£ĪļÅä ņ┤ēņ¦äņŗ£Ēéżļ®░ ĻĖ░ļŖźņä▒ ļ¼╝ņ¦łļōżņØä ņāØņä▒ĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż(Andrews, 1992; Xu ļō▒ 2016). ĒĢśņ¦Ćļ¦ī ļ»ĖņāØļ¼╝ņØś ĒÖ£ņä▒ņØ┤ ņØ╝ņĀĢĒĢśņ¦Ć ļ¬╗ĒĢśļŗżļŖö ļŗ©ņĀÉņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā, ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ļŖö ļåŹņŚģ ņāØĒā£Ļ│äņŚÉ ĒåĀņ¢æ ļ│æĒĢ┤ļź╝ Ļ░ÉņåīĒĢĀ ņłś ņ׳ļŖö ļīĆņĢłņ£╝ļĪ£ ņŚ¼Ļ▓©ņ¦äļŗż(WhippsņÖĆ McQuilken, 2009). ļö░ļØ╝ņä£ ņĄ£ĻĘ╝ ĒŖ╣ņĀĢĒĢ£ ņ¦ĆņŚŁņØ┤ļéś ĻĘ╣ĒĢ£ ņä£ņŗØ ĒÖśĻ▓ĮņŚÉņä£ ņāØļ¼╝ĒÖ£ņä▒ņØ┤ ņÜ░ņłśĒĢ£ ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØś ĒāÉņāēņØ┤ ņØ┤ļŻ©ņ¢┤ņ¦ĆĻ│Ā ņ׳ļŗż(Dharni ļō▒, 2012; Kavamura ļō▒, 2013).
ņןņłśĒÆŹļÄģņØ┤(Allomyrina dichotoma L.)ļŖö ļö▒ņĀĢļ▓īļĀłļ¬®(Coleoptera) ĒÆŹļÄģņØ┤Ļ│╝(Scarabaeidae)ņŚÉ ņåŹĒĢśļŖö Ļ│żņČ®ņ£╝ļĪ£ ĒĢ£ĻĄŁ, ņØ╝ļ│Ė, ņżæĻĄŁ, ņØĖļÅä ļō▒ņŚÉņä£ ļČäĒżĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĪīļŗż. ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØĆ ļ¼┤ĻĖ░ņ¦ł, ņä¼ņ£Āņ¦ł, ļ»ĖņāØļ¼╝ ļō▒ņØ┤ ĒżĒĢ©ļÉ£ ļČĆņŚĮĒåĀ ļ░Å ļČĆņŗØĒåĀļź╝ ļ©╣Ļ│Ā ņ×ÉļØ╝ļ®░, ņ×æļ¼╝ņØä Ļ░ĆĒĢ┤ĒĢśņ¦Ć ņĢŖņĢä ļ╣äĻĄÉņĀü ĒåĀņ¢æĻ│╝ ņ╣£ĒÖöņĀüņØ┤ļŗż. ļśÉĒĢ£, ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØĆ ļ»╝Ļ░äņĢĮņĀ£ļĪ£ņä£ Ļ▒┤Ļ░Ģļ│┤ņĪ░ņÜ® ņĢĮņĀ£ļĪ£ ļäÉļ”¼ ņØ┤ņÜ®ļÉśĻ│Ā ņ׳ņ£╝ļ®░, Ļ░ä ņ¦łĒÖś, ņä▒ņØĖļ│æ ņ╣śļŻī ļ░Å ņśłļ░®ņŚÉ ļīĆĒĢ£ ĒÜ©Ļ│╝Ļ░Ć ļ│┤Ļ│ĀļÉśņŚłļŗż(Choi ļō▒, 2006; Chung ļō▒, 2013).
ņØ┤ļ¤¼ĒĢ£ Ļ┤ĆņĀÉņŚÉņä£ Ļ│ĀĒÜ©ņ£©ņØś ĒÜ©ņåīļź╝ ņāØņé░ĒĢśļŖö Ļ│żņČ® ņןļé┤ļ»ĖņāØļ¼╝ņØĆ ņŗØļ¼╝ ĻĘĀļźśņØś ņÖĖļ¦ēņØä Ļ░ĆņłśļČäĒĢ┤ĒĢĀ ņłś ņ׳ļŖö cellulase, protease, chitinaseņÖĆ Ļ░ÖņØĆ ĒÜ©ņåī ĒÖ£ņä▒ņØ┤ ļø░ņ¢┤ļéĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉśļ®░, ļśÉĒĢ£ Ļ│żņČ®ņØś ņåīĒÖöĻĖ░Ļ┤Ćļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ĒĢäņłśņĀüņØĖ ņĢäļ»ĖļģĖņé░ņØ┤ļéś ļ╣äĒāĆļ»╝ ļō▒ņØä ĒĢ®ņä▒ĒĢśļŖö Ļ│ĄņāØņ×ÉņØĖ ņןļé┤ ļ»ĖņāØļ¼╝ņØ┤ ņŗØļ¼╝ņØś ņāØņ£ĪņØ┤ļéś ņāØņןņŚÉļÅä ļ░ĆņĀæĒĢ£ Ļ┤ĆĻ│äļź╝ Ļ░Ćņ¦ĆĻ│Ā ņ׳ņØä Ļ░ĆļŖźņä▒ņØ┤ ļåÆļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż.
ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ļŖö ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ņåīĒÖöĻĖ░Ļ┤Ćņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ļÉ£ ņןļé┤ ļ»ĖņāØļ¼╝ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ļŗżņ¢æĒĢ£ ņŗØļ¼╝ ĻĘĀļźśļ│æņŚÉ ļīĆĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒Ļ│╝ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ Ļ▓ĆņĀĢĒĢśņśĆĻ│Ā, ņŗØļ¼╝ņØś ņāØņ£ĪĻ│╝ ļ│æ ļ░®ņĀ£ņŚÉ ņ¦üŌĆóĻ░äņĀæņĀüņ£╝ļĪ£ Ļ┤ĆņŚ¼ĒĢśļŖö ņ×æņÜ®ļ®öņ╗żļŗłņ”śĻ│╝ ņŗØļ¼╝ņāØņןņ┤ēņ¦äĒÖ£ņä▒ņØä ĒÅēĻ░ĆĒĢśņśĆļŗż.
ņ×¼ļŻī ļ░Å ļ░®ļ▓Ģ
ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ® ņ▒äņ¦æ ļ░Å ņןļé┤ ļ»ĖņāØļ¼╝ ļČäļ”¼
ņŗØļ¼╝ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä Ļ░Ćņ¦ĆļŖö ņןļé┤ ļ»ĖņāØļ¼╝ņØś ĒāÉņāēņŚÉ ņØ┤ņÜ®ļÉ£ ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØĆ Ļ░ĢņøÉļÅä Ļ░Ģļ”ēņŗ£ ņØĖĻĘ╝ ņ×äņé░ ļČĆņé░ļ¼╝ņŚÉņä£ ņ▒äņ¦æĒĢ£ 3ļĀ╣ ņ£ĀņČ®ņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. ņןņłśĒÆŹļÄģņØ┤ 3ļĀ╣ ņ£ĀņČ®ņØä ļ®ĖĻĘĀļÉ£ 70% ņŚÉĒāäņś¼ļĪ£ 1ŌĆÆ2ļČä ļÅÖņĢł ņ╣©ņ¦ĆĒĢśņŚ¼ ņ┤Øņ▓┤ Ēæ£ļ®┤ņØś ņśżņŚ╝ņøÉņØä ņĀ£Ļ▒░ĒĢ£ Ēøä, ļ®ĖĻĘĀņłśļĪ£ 2ĒÜī ņäĖņ▓ÖĒĢśņśĆļŗż. ĒĢ┤ļČĆņäĖĒŖĖļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņןņłśĒÆŹļÄģņØ┤ 3ļĀ╣ ņ£ĀņČ®ņØś ļ¬ĖĒåĄ ļČĆļČäņŚÉņä£ ļ░░ļź╝ ņĀłĻ░£ĒĢśņŚ¼ ņ▓┤Ēæ£ ļČĆļČäņØä ņĀ£ņÖĖĒĢ£ ņןņØä ņĘ©ĒĢśņŚ¼ ņĀäņן(foregut), ņżæņן(midgut), Ēøäņן(hindgut)ņ£╝ļĪ£ ņŚ░Ļ▓░ļÉ£ ņåīĒÖöĻĖ░Ļ┤ĆņØä ļ¼┤ĻĘĀņĀüņ£╝ļĪ£ ļČäļ”¼ĒĢśņśĆļŗż. Ļ░üĻ░ü ļČäļ”¼ļÉ£ ņåīĒÖöĻĖ░Ļ┤ĆņØś ņŗ£ļŻīļź╝ 1 gņØä ņ▒äņĘ©ĒĢśņŚ¼ ļ®ĖĻĘĀņłś 9 mlņŚÉ ļäŻĻ│Ā ņČ®ļČäĒ׳ ĻĘĀņ¦łĒÖöĒĢ£ Ēøä, ļŗ©Ļ│äņĀüņ£╝ļĪ£ ĒؼņäØĒĢśņŚ¼ potato dextrose agar (PDA), tryptic soy agar, starch casein agar ļ░░ņ¦ĆņŚÉ Ļ░üĻ░ü ļÅäļ¦ÉĒĢśņŚ¼ 25┬░CņŚÉņä£ 3-10ņØ╝Ļ░ä ļ░░ņ¢æĒĢśļ®┤ņä£ ĒÅēĒīÉļ░░ņ¦ĆņŚÉ ĒśĢņä▒ļÉ£ colonyļź╝ ņł£ņłś ļČäļ”¼ĒĢśņśĆļŗż.
ĒĢŁĻĘĀĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņł£ņłś ļČäļ”¼ļÉ£ ņןļé┤ ļ»ĖņāØļ¼╝ļōżņØä ļīĆņāüņ£╝ļĪ£ ņŗØļ¼╝ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢśĻ│Āņ×É 8ņóģņØś ņŗØļ¼╝ļ│æņøÉĻĘĀ ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ(Botrytis cinerea), ĒåĀļ¦łĒåĀ ņŗ£ļōżņØīļ│æ(Fusarium. oxysporum f. sp. radicis-lycopersici), ļ▓╝ ļÅäņŚ┤ļ│æ(Magnaporthe grisea), ļ▓╝ ņ×Äņ¦æļ¼┤ļŖ¼ļ¦łļ”äļ│æ(Rhizoctonia solani AG-1(1A)), Ļ│ĀņČö ĒāäņĀĆļ│æ(Colletotrichum acutatum), Ļ│ĀņČö ņŚŁļ│æ(Phytophthora capsici), ļ¦łļŖś ĒØæņāēņŹ®ņØīĻĘĀĒĢĄļ│æ(Sclerotium cepivorum), ļ░░ņČö ļ░æļæźņŹ®ņØīļ│æ(R. solani)Ļ│╝ PDA ļ░░ņ¦Ć ņāüņŚÉņä£ ļīĆņ╣śļ░░ņ¢æ(dual culture)ĒĢś ņŚ¼ 25┬░CņŚÉņä£ 10ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ļ│æņøÉĻĘĀņØś ĻĘĀņé¼ņ▓┤ņØś ņāØņןņ¢ĄņĀ£ ņĀĢļÅä (inhibition zone)ļź╝ ņĪ░ņé¼ĒĢśņŚ¼ ĒśĢņä▒ļÉ£ clear zone (mm)ņØś ĻĖĖņØ┤ļĪ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ņé¼ņÜ®ļÉ£ ņŗØļ¼╝ļ│æņøÉĻĘĀņØĆ Ļ░Ģļ”ēņøÉņŻ╝ļīĆĒĢÖĻĄÉ ņŗØļ¼╝ļ│æļ”¼ņŗżĒŚśņŗżņŚÉņä£ ļČäļ”¼ ļÅÖņĀĢĒĢśņŚ¼ ņé¼ņÜ®ĒĢśĻ▒░ļéś, ļåŹņŚģņ£ĀņĀäņ×ÉņøÉ ņä╝Ēä░ņŚÉņä£ ļČäņ¢æ ļ░øņĢä ņé¼ņÜ®ĒĢśņśĆļŗż.
ĻĖĖĒĢŁ ļ»ĖņāØļ¼╝ņØś ņāØļ¼╝ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ļīĆņ╣śļ░░ņ¢æņØä ĒåĄĒĢ┤ ņäĀļ░£ļÉ£ 38ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ņØś ņāØļ¼╝ĒÖ£ņä▒ņØĆ ņś©ņŗżņŚÉņä£ ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ, ļ░░ņČö ļ┐īļ”¼Ēś╣ļ│æ(Plasmodiophora brassicae), Ļ│ĀņČö ĒāäņĀĆļ│æ, Ļ│ĀņČö ņŚŁļ│æņØä ļīĆņāüņ£╝ļĪ£ ņłśĒ¢ēĒĢśņśĆļŗż. ņŗØļ¼╝ ņ×¼ļŻī ņżĆļ╣äļŖö ĒåĀļ¦łĒåĀ(Superdotaerang, Koregon, Anseong, Korea), Ļ│ĀņČö(Supermanita, Nongwoobio, Suwon, Korea), ļ░░ņČö(Chunkwang, Sakata Korea, Seoul, Korea)ņØś Ļ░üĻ░üņØś ņóģņ×Éļź╝ ņøÉņśłņÜ®ņāüĒåĀ(Chologi, Nongwoobio)ļĪ£ ņ▒äņøīņ¦ä 162 plug traysņŚÉ ĒīīņóģĒĢśņŚ¼ 3ņŻ╝ ļÅÖņĢł ņ£Īļ¼śĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆĻ│Ā, 3ņŻ╝ļÉ£ ņ£Īļ¼śļź╝ ĒżĒŖĖ(10 cm diameter)ņŚÉ ņØ┤ņŗØĒĢśņŚ¼ ĒÖ£ņ░®ņŗ£Ēé© ņ£Īļ¼śļź╝ ņé¼ņÜ®ĒĢśņśĆļŗż. ĻĖĖĒĢŁņäĖĻĘĀ ĒśäĒāüņĢĪ ņżĆļ╣äļź╝ ņ£äĒĢ┤ ņØ╝ļ░ś ņäĖĻĘĀņØĆ tryptic soy brothņŚÉ 180 rpmņ£╝ļĪ£ 28┬░C, 24ņŗ£Ļ░ä ļÅÖņĢł ļ░░ņ¢æĒĢśņŚ¼ 1.0├Ś108ŌĆÆ109 cfu/ml ņĪ░ņĀĢĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆĻ│Ā, ļ░®ņäĀĻĘĀņØĆ starch casein broth ļ░░ņ¦ĆņŚÉ 180 rpmņ£╝ļĪ£ 25┬░C, 7ņØ╝ ļÅÖņĢł ļ░░ņ¢æĒĢ£ ļ░░ņ¢æņĢĪņØä ņé¼ņÜ®ĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ Ļ▓ĆņĀĢņØä ņ£äĒĢ┤ B. cinerea ĻĘĀņŻ╝ļź╝ ļ¬ģņĪ░Ļ▒┤ņŚÉņä£ 3ņŻ╝Ļ░ä ļ░░ņ¢æĒĢśņŚ¼ 0.5% glucose, 0.25% K2HPO4 ĒśäĒāüņĢĪņ£╝ļĪ£ ņłśĒÖĢĒĢśņŚ¼ Ēżņ×É ļåŹļÅäļź╝ 1.0├Ś106 spore/mlļĪ£ ņĪ░ņĀĢĒĢśņśĆļŗż. ĻĖĖĒĢŁĻĘĀ ĒśäĒāüņĢĪņØä ĒåĀļ¦łĒåĀ ņ×ÄņŚÉ ņČ®ļČäĒ׳ ņĀüņŗ£ļÅäļĪØ ļČäļ¼┤ņ▓śļ”¼ĒĢśņśĆĻ│Ā, 24ņŗ£Ļ░ä ĒøäņŚÉ Ēżņ×É ĒśäĒāüņĢĪņØä ļČäļ¼┤ ņĀæņóģĒĢ£ Ēøä Ļ│ĀņŖĄļÅä ņŖĄņŗżņāü(95% relative humidity [RH], 20┬░C)ņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ļ│æ ļ░£ņāØņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░£ļ│æļÅäļŖö 5ļŗ©Ļ│ä(0, ļ░£ļ│æĒĢśņ¦Ć ņĢŖņØī; 1, 0.1ŌĆÆ5% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 2, 5.1ŌĆÆ20% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 3, 20.1ŌĆÆ40% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 4, 40.1ŌĆÆ100% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ)ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░░ņČö ļ┐īļ”¼Ēś╣ļ│æ Ļ▓ĆņĀĢņØä ņ£äĒĢ┤ ņä▒ņłÖĒĢ£ ļ┐īļ”¼Ēś╣ņØä ņ▒äņ¦æĒĢśņŚ¼ blenderļĪ£ ļ¦łņćäĒĢśņŚ¼ ĒśäĒāüņĢĪņØä ļ¦īļōĀ Ēøä ņøÉņśłņÜ®ņāüĒåĀ(Chologi, Nongwoobio)ņÖĆ ņä×ņ¢┤ņä£ ņśżņŚ╝ĒåĀļź╝ ļ¦īļōżņŚłļŗż(5 l/20 kg). ļ░░ņČö ļ┐īļ”¼Ēś╣ļ│æ ņśżņŚ╝ĒåĀļź╝ ĒżĒŖĖ(10 cm diameter)ņŚÉ ļäŻĻ│Ā, ĻĖĖĒĢŁĻĘĀ ĒśäĒāüņĢĪņØ┤ ņ▓śļ”¼ļÉ£ ļ░░ņČö ņ£Āļ¼śļź╝ ņØ┤ņŗØ Ēøä 40ņØ╝Ļ░ä ņ£Īļ¼śĒĢ£ Ēøä ļ│æ ļ░£ņāØņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░£ļ│æļÅäļŖö 5ļŗ©Ļ│ä(0, ļ┐īļ”¼Ēś╣ ņŚåņØī; 1, 1ŌĆÆ9% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 2, 10ŌĆÆ30% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 3, 31ŌĆÆ60% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 4, 60ŌĆÆ100% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ)ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż. Ļ│ĀņČö ņŚŁļ│æ Ļ▓ĆņĀĢņØä ņ£äĒĢ┤ P. capsici ĻĘĀņŻ╝ļź╝ oatmeal agarņŚÉ ņĀæņóģĒĢśņŚ¼ 25┬░C, ļ¬ģņĪ░Ļ▒┤ņŚÉņä£ 10ņØ╝Ļ░ä ļ░░ņ¢æĒĢśņśĆļŗż. ļ░░ņ¦Ć Ēæ£ļ®┤ņŚÉ ņāØņä▒ļÉ£ ņŚŁļ│æĻĘĀņØä 4┬░C ņ░©Ļ░ĆņÜ┤ ļ®ĖĻĘĀņłśļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņ£ĀņŻ╝ņ×ÉļéŁņØä ĒÜīņłśĒĢśņŚ¼ ļåŹļÅäļź╝ 1.0├Ś103 zoosporangia/ml ņĪ░ņĀĢ Ēøä ņ£ĀņŻ╝ņ×ÉĻ░Ć ļ░®ņČ£ļÉśļÅäļĪØ ņ£ĀļÅäĒĢśņśĆļŗż. Ļ░üĻ░üņØś ĒżĒŖĖņŚÉ ĻĖĖĒĢŁĻĘĀ ĒśäĒāüņĢĪ 20 mlņö® Ļ┤ĆņŻ╝ņ▓śļ”¼ 24ņŗ£Ļ░ä ĒøäņŚÉ ņ£ĀņŻ╝Ēżņ×É ĒśäĒāüņĢĪņØä 10 mlņö® Ļ┤ĆņŻ╝ņ▓śļ”¼ĒĢśņŚ¼ Ļ│ĀņŖĄļÅä ņŖĄņŗżņāü(95% RH, 25┬░C)ņŚÉņä£ 24ņŗ£Ļ░ä ļÅÖņĢł ņŖĄņŗż ņ▓śļ”¼ĒĢśĻ│Ā 10ņØ╝ ļÅÖņĢł ļ░£ļ│æ ņŗ£Ēé© Ēøä ļ░£ļ│æļÅäļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░£ļ│æļÅäļŖö 6ļŗ©Ļ│ä(0, ļ░£ļ│æĒĢśņ¦Ć ņĢŖņØī; 1, ņ¦ĆņĀ£ļČĆĻ░Ć ļ¼┤ļź┤Ļ│Ā Ļ░łļ│ĆĒĢśĻĖ░ ņŗ£ņ×æ; 2, 30ŌĆÆ50% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 3, 50ŌĆÆ70% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 4, 70ŌĆÆ90% ņĀĢļÅäņØś ļ│æ ļ░£ņāØ; 5, Ļ│Āņé¼)ļĪ£ ņĪ░ņé¼ĒĢśņśĆļŗż. Ļ│ĀņČö ĒāäņĀĆļ│æ Ļ▓ĆņĀĢņØä ņ£äĒĢ┤ C. acutatum ĻĘĀņŻ╝ļź╝ PDA ļ░░ņ¦ĆņŚÉ ņĀæņóģĒĢśņŚ¼ 25┬░C, 14ņØ╝ ļÅÖņĢł ļ¬ģņĪ░Ļ▒┤ņŚÉņä£ ļ░░ņ¢æĒĢśņŚ¼ Ēżņ×É ĒśĢņä▒ņØä ņ£ĀļÅäĒĢśņśĆļŗż. ļ░░ņ¦Ć Ēæ£ļ®┤ņŚÉ ĒśĢņä▒ļÉ£ ĒāäņĀĆļ│æĻĘĀņØä ļ®ĖĻĘĀņłśļĪ£ ĒÜīņłśĒĢśņŚ¼ 1.0├Ś105 spore/mlņØ┤ ļÉśļÅäļĪØ ņĪ░ņĀĢĒĢśņŚ¼ ņé¼ņÜ®ĒĢśņśĆļŗż. ņĀĢņŗØ Ēøä 10ņŻ╝ļÉ£ Ļ│ĀņČö(Supermanita, Nongwoobio)ņŚÉņä£ ņłśĒÖĢĒĢ£ ļ╣©Ļ░ä Ļ│╝ņŗżņØä 1% sodium hypochlorite solution (NaOCl)ņŚÉ Ēæ£ļ®┤ ņé┤ĻĘĀĒĢ£ Ēøä ļ®ĖĻĘĀņłśņŚÉ 2ĒÜī ņäĖņ▓ÖĒĢśņśĆļŗż. ņśżņŚ╝ņøÉņØ┤ ņĀ£Ļ▒░ļÉ£ Ļ│ĀņČö Ļ│╝ņŗżņØä Ļ▒┤ņĪ░ Ēøä ļ®ĖĻĘĀļÉ£ ļ░öļŖśņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņØĖņ£äņĀüņ£╝ļĪ£ ņāüņ▓śļź╝ ļéĖ Ēøä ĻĖĖĒĢŁņäĖĻĘĀ ĒśäĒāüņĢĪņØä 20 ╬╝lņö® pipettingĒĢśņśĆļŗż. ĻĖĖĒĢŁĻĘĀ ĒśäĒāüņĢĪ ņ▓śļ”¼ 24ņŗ£Ļ░ä ĒøäņŚÉ ĒāäņĀĆļ│æĻĘĀ Ēżņ×É ĒśäĒāüņĢĪ 10 ╬╝lņö® pipettingĒĢ£ Ēøä Ļ│ĀņŖĄļÅä ņŖĄņŗżņāü(95% RH, 20┬░C)ņŚÉņä£ 5ņØ╝Ļ░ä ļ░░ņ¢æĒĢ£ Ēøä ļ│æļ░ś ļČĆņ£äļź╝ diameter (mm) ĻĖĖņØ┤ļź╝ ņĪ░ņé¼ĒĢśņśĆļŗż. ļ░®ņĀ£Ļ░Ć (%)ļŖö ņĢäļל Ļ│ĄņŗØņŚÉ ļö░ļØ╝ ņé░ņČ£ĒĢśņśĆļŗż. ļ░®ņĀ£Ļ░Ć(%)=([ļ¼┤ņ▓śļ”¼ĻĄ¼ņØś ļ│æļ░ś diameter (mm)ŌĆÆņ▓śļ”¼ĻĄ¼ņØś ļ│æļ░ś diameter (mm)]/ļ¼┤ņ▓śļ”¼ĻĄ¼ ņØś ļ│æļ░ś diameter (mm))├Ś100.
ņäĀļ░£ ĻĘĀņŻ╝ņØś ļÅÖņĀĢ
ņāØļ¼╝ĒÖ£ņä▒ Ļ▓ĆņĀĢņØä ĒåĄĒĢ┤ ņĄ£ņóģ ņäĀļ░£ļÉ£ 10ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ņØä ļÅÖņĀĢĒĢśĻĖ░ ņ£äĒĢ┤ ņŻ╝ņé¼ņĀäņ×ÉĒśäļ»ĖĻ▓ĮņØä ĒåĄĒĢ┤ ĒśĢĒā£ĒĢÖņĀü ĒŖ╣ņ¦ĢņØä Ļ▓ĆņĀĢĒĢśņśĆĻ│Ā, 16S rRNA gene ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢśņśĆļŗż. ņäĀļ░£ļÉ£ ĻĘĀņŻ╝ņØś total genomic DNAņØś ņČöņČ£ņØĆ Nishiguchi ļō▒(2002)ņØś ļ░®ļ▓ĢņØä ņØ┤ņÜ®ĒĢśņśĆļŗż. ņČöņČ£ļÉ£ genomic DNAļŖö universal primer 27F (5ŌĆ▓-AGAGTTTGATCMTGGCTCAG-3ŌĆ▓)ņÖĆ 1492R (5ŌĆ▓-TACGGYTACCTTGTTACGACTT-3ŌĆ▓) ĒöäļØ╝ņØ┤ļ©Ėļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ PCR ņ”ØĒÅŁĒĢśņśĆļŗż. ņ”ØĒÅŁļÉ£ PCR ņé░ļ¼╝ņØä ņĀĢņĀ£ĒĢśņŚ¼, Solgent Co. Ltd. (Seoul, Korea)ņŚÉ ņØśļó░ĒĢśņŚ¼ 16S rRNA gene ņŚ╝ĻĖ░ņä£ņŚ┤ņØä Ļ▓░ņĀĢĒĢśņśĆĻ│Ā, Ļ▓░ņĀĢļÉ£ ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ SeqMan (DNASTAR, Madison, WI, USA)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ņĀĢļ”¼ĒĢśņśĆļŗż. 16S rRNA gene ņŚ╝ĻĖ░ņä£ņŚ┤ņØĆ NCBI (National Center for Biotechnology Information) BlastN databaseļź╝ ĒåĄĒĢśņŚ¼ ņāüļÅÖņä▒ņØä ļ╣äĻĄÉĒĢśņśĆļŗż. MEGA 6ņØś Cluster Wļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ņĀĢļĀ¼ĒĢśņŚ¼ neighbor-joining ļ░®ļ▓ĢņŚÉ ļö░ļØ╝ Ļ│äĒåĄļÅäļź╝ ņ×æņä▒ĒĢśņśĆĻ│Ā, bootstrappingņØĆ 1,000ĒÜī ļ░śļ│ĄĒĢśņśĆļŗż.
ņäĀļ░£ ĻĘĀņŻ╝ņØś IAA, ņØĖņé░ Ļ░ĆņÜ®ĒÖö, siderophore ņāØņä▒ ĒÖ£ņä▒, Ļ░ĆņłśļČäĒĢ┤ ĒÜ©ņåī Ļ▓ĆņĀĢ
IAA (indole-3-acetic acid) ņāØņä▒ĒÖ£ņä▒ņØĆ IAAņØś ņĀäĻĄ¼ļ¼╝ņ¦łņØĖ L-tryptophan 0.1% ņ▓©Ļ░ĆĒĢ£ KingŌĆÖs B broth ļ░░ņ¦ĆņŚÉ ļ░░ņ¢æļÉ£ ļ░░ņ¢æņĢĪĻ│╝ Salkowsky reagent (35% perchloric acid 50 ml, 0.5 M ferric chloride 1 ml)ļź╝ 1:2 (v/v)ļĪ£ Ēś╝ĒĢ®ĒĢśņŚ¼ ļ░śņØæņŗ£ņ╝£ ĒÖĢņØĖ Ēøä 530 nmņØś ĒīīņןņŚÉņä£ ĒØĪĻ┤æļÅäļź╝ ņĖĪņĀĢĒĢśņśĆļŗż. ņØĖņé░ Ļ░ĆņÜ®ĒÖö(phosphate solubilizing) ĒÖ£ņä▒ņØĆ PikovskayaŌĆÖs agar [glucose 10.0 g, (NH4)2SO4 0.5 g, KCl 0.2 g, MgSO4 0.1 g, MnSO4 0.002 g, FeSO4 0.002 g, yeast extract 0.5 g, Ca3(PO4)2 5 g, agar 15 g, distilled water 1 l] ļ░░ņ¦Ć(Zaidi ļō▒, 2006)ņŚÉņä£ Ļ▓ĆņĀĢĒĢśņśĆņ£╝ļ®░, Ēł¼ļ¬ģĒÖś(clear zone)ņØä ĒśĢņä▒ĒĢśļŖö Ļ▓āņØ┤ ĒÖ£ņä▒ņØ┤ ņ׳ļŖö Ļ▓āņ£╝ļĪ£ Ļ░äņŻ╝ĒĢśņśĆļŗż. Siderophore ņāØņä▒ĒÖ£ņä▒ņØĆ Chrome azourol S (CAS) agar ļ░░ņ¦Ć(SchwynņÖĆ Neilands, 1997)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ Ļ▓ĆņĀĢĒĢśņśĆļŗż. CAS ĒÅēĒīÉļ░░ņ¦ĆņŚÉ 5 mmņØś cork borrerļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ ļ░░ņ¦Ć ņżæņĢÖņŚÉ discļź╝ ņĀ£Ļ▒░ĒĢ£ Ēøä, ņäĀļ░£ļÉ£ ĻĘĀņŻ╝ ļ░░ņ¢æņĢĪņØä 20 ╬╝lļź╝ pipettingĒĢśņŚ¼ ņĀæņóģĒĢśņśĆļŗż. ņĀæņóģ Ēøä 28┬░CņŚÉņä£ 14ņØ╝ ļÅÖņĢł ļ░░ņ¢æĒĢśļ®┤ņä£ orange halo zoneņØś ĒśĢņä▒ ņ£Āļ¼┤ļĪ£ siderophoreņØś ĒÖ£ņä▒ņØä ņĪ░ņé¼ĒĢśņśĆļŗż. ņŗØļ¼╝ļ│æņøÉĻĘĀņØś ņäĖĒżļ▓ĮņØä ļČäĒĢ┤ĒĢśļŖö ĒÜ©ņåī ĒÖ£ņä▒ņØĆ ╬▓-1,3-glucanase ĒÖ£ņä▒Ļ│╝ protease ĒÖ£ņä▒ ĒåĄĒĢ┤ Ļ▓ĆņĀĢĒĢśņśĆļŗż. ╬▓-1,3-glucanase ĒÖ£ņä▒ņØĆ 1% carboxymethyl-celluloseļź╝ ĒĢ©ņ£ĀĒĢ£ casein-yeast extract agar (casein 5 g, yeast extract 2.5 g, glucose 1 g, agar 18 g, distilled water 1 l)ļ░░ņ¦ĆņŚÉņä£ ļ░░ņ¢æĒĢśņŚ¼ Congo red ņÜ®ņĢĪ(1 mg/ml)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ 15ļČä ļÅÖņĢł ņŚ╝ņāē Ēøä 1.0 M NaClļĪ£ 3ļ▓ł ņäĖņ▓ÖĒĢśņŚ¼ Ēł¼ļ¬ģĒÖś ĒśĢņä▒ņØä diameter (mm)ļĪ£ ĒÖ£ņä▒ ļŖźļĀźņØä ņĪ░ņé¼ĒĢśņśĆļŗż. Protease ĒÖ£ņä▒ņØĆ protease ĒÖ£ņä▒ ļ░░ņ¦Ć(skim milk [Sigma Aldrich Co., St. Louis, MO, USA] 10 g, nutrient broth [Becton, Dickinson and Company, Sparks, MD, USA] 2 g, agar 15 g, distilled water 1 l) ļ░░ņ¦ĆņŚÉ ļ░░ņ¢æĒĢśņŚ¼ Ēł¼ļ¬ģĒÖś ĒśĢņä▒ņØä diameter (mm)ļĪ£ ĒÖ£ņä▒ ļŖźļĀźņØä ņĪ░ņé¼ĒĢśņśĆļŗż.
ņŗØļ¼╝ņāØņןņ┤ēņ¦äĒÖ£ņä▒ Ļ▓ĆņĀĢ
3ņŻ╝ ļÅÖņĢł ĒéżņÜ┤ ĒåĀļ¦łĒåĀ, Ļ│ĀņČö ņ£Āļ¼śļź╝ ĒżĒŖĖ(15├Ś15 cm diameter)ņŚÉ ņØ┤ņŗØ Ēøä ĻĖĖĒĢŁĻĘĀ ĒśäĒāüņĢĪņØä 100 mlņö® 7ņØ╝ Ļ░äĻ▓®ņ£╝ļĪ£ 3ĒÜī Ļ┤ĆņŻ╝ ņ▓śļ”¼ĒĢśņśĆļŗż. ļ¦łņ¦Ćļ¦ē ņ▓śļ”¼ 7ņØ╝ Ēøä, ņżäĻĖ░ ņ¦üĻ▓Į, ĻĖĖņØ┤, ņāØņ▓┤ ņżæļ¤ē, Ļ▒┤ņĪ░ ņżæļ¤ēĻ│╝ ļ┐īļ”¼ ĻĖĖņØ┤, ņāØņ▓┤ ņżæļ¤ē, Ļ▒┤ņĪ░ ņżæļ¤ē ĻĘĖļ”¼Ļ│Ā ņŚĮļĪØņåī ĒĢ©ļ¤ēņØä ņĖĪņĀĢĒĢśņśĆļŗż.
ĒåĄĻ│äļČäņäØ
ĒĢŁĻĘĀĒÖ£ņä▒Ļ│╝ ĒÜ©ņåīĒÖ£ņä▒ Ļ▓ĆņĀĢņØĆ 3ļ░śļ│Ą 3ĒÜī ņłśĒ¢ēĒĢśņśĆņ£╝ļ®░, ņāØļ¼╝ĒÖ£ņä▒Ļ│╝ ņāØņןņ┤ēņ¦äĒÖ£ņä▒ Ļ▓ĆņĀĢņØĆ ņ▓śļ”¼ĻĄ¼ļŗ╣ 5ĒżĒŖĖņö® 3ļ░śļ│Ą ņÖäņĀäņ×äņØśļ░░ņ╣śļ▓Ģņ£╝ļĪ£ ņłśĒ¢ēĒĢśņśĆļŗż. ņŗżĒŚśņ£╝ļĪ£ ņ¢╗ņ¢┤ņ¦ä Ļ▓░Ļ│╝ļŖö SAS system (SAS Institute Inc., Cary, NC, USA)ņØä ņØ┤ņÜ®ĒĢśņŚ¼ ĒåĄĻ│äļČäņäØņØä ĒĢśņśĆņ£╝ļ®░, DuncanņØś ļŗżņżæĻ▓ĆņĀĢ(DuncanŌĆÖs multiple range test)ļź╝ ņØ┤ņÜ®ĒĢśņŚ¼ PŌēż0.05 ņłśņżĆņŚÉņä£ Ļ░ü ņ▓śļ”¼ Ļ░äņØś ņ£ĀņØśņä▒ņØä Ļ▓Ćņ”ØĒĢśņśĆļŗż.
Ļ▓░Ļ│╝ ļ░Å Ļ│Āņ░░
ņןļé┤ņäĖĻĘĀ ļČäļ”¼ ļ░Å ĒĢŁĻĘĀĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ņåīĒÖöĻĖ░Ļ┤Ć ņĀäņן, ņżæņן, ĒøäņןņŚÉņä£ ņ┤Ø 265ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ņØä ļČäļ”¼ĒĢśņśĆĻ│Ā, ļČäļ”¼ĒĢ£ ļ»ĖņāØļ¼╝ņØä 8ņóģņØś ņŗØļ¼╝ļ│æņøÉĻĘĀ ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ, ĒåĀļ¦łĒåĀ ņŗ£ļōżņØīļ│æ, ļ▓╝ ļÅäņŚ┤ļ│æ, ļ▓╝ ņ×Äņ¦æļ¼┤ļŖ¼ļ¦łļ”äļ│æ(R. solani AG-1(1A)), Ļ│ĀņČö ĒāäņĀĆļ│æ, Ļ│ĀņČö ņŚŁļ│æ, ļ¦łļŖś ĒØæņāēņŹ®ņØīĻĘĀĒĢĄļ│æ, ļ░░ņČö ļ░æļæźņŹ®ņØīļ│æņŚÉ ļīĆĒĢśņŚ¼ ļīĆņ╣śļ░░ņ¢æ Ļ▓░Ļ│╝, ņ┤Ø 148ņóģņØś ĻĖĖĒĢŁĻĘĀņØä ņŖżĒü¼ļ”¼ļŗØĒĢśņśĆļŗż. ņĀäņןņŚÉņä£ 33ņóģņ£╝ļĪ£ Ļ░Ćņן ļé«ņØĆ ĻĖĖĒĢŁĻĘĀņØś ļČäļ”¼ļ╣äļź╝ ļ│┤ņśĆĻ│Ā, ņżæņןņŚÉņä£ 52ņóģ, ĒøäņןņŚÉņä£ 63ņóģņ£╝ļĪ£ Ļ░Ćņן ļ¦ÄņØĆ ĻĖĖĒĢŁĻĘĀņØś ļČäļ”¼ļ╣äļź╝ ļéśĒāĆļāłļŗż. ņØ┤ļ¤¼ĒĢ£ ĻĖĖĒĢŁĻĘĀņØś ļČäļ”¼ļ╣äļŖö Ļ│żņČ®ņØś ņäŁņŗØĻĄ¼ņĪ░ņÖĆ ņåīĒÖöĻĖ░Ļ┤ĆņŚÉ ļö░ļźĖ ĒŖ╣ņ¦Ģņ£╝ļĪ£ ļ│┤ņŚ¼ņ¦äļŗż. ļśÉĒĢ£, 8ņóģņØś ņŗØļ¼╝ļ│æņøÉĻĘĀ ņżæ F. oxysporum f. sp. radicis-lycopersiciņÖĆ R. solaniņŚÉ ļīĆĒĢ£ ĒÖ£ņä▒ņØä ņ¦Ćļŗī ĻĖĖĒĢŁĻĘĀņØś ļČäļ”¼ļ╣äļŖö ļé«Ļ▓ī ļéśĒāĆļé¼Ļ│Ā, S. cepivorumņÖĆ M. griseaņŚÉ ĒÖ£ņä▒ņØä Ļ░Ćņ¦ä ĻĖĖĒĢŁĻĘĀņØś ļČäļ”¼ļ╣äļŖö ļåÆĻ▓ī ļéśĒāĆļé¼ļŗż. ļīĆņ╣śļ░░ņ¢æņØä ĒåĄĒĢ┤ ņäĀļ░£ļÉ£ 38ņóģņØś ĻĖĖĒĢŁĻĘĀņØĆ ļīĆļČĆļČäņØś ņŗØļ¼╝ ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä ļ│┤ņŚ¼, ņŖżĒÄÖĒŖĖļ¤╝ņØ┤ ļäōņØĆ Ļ▓āņØä ĒÖĢņØĖĒĢśņśĆļŗż(Table 1).
ĻĖĖĒĢŁ ļ»ĖņāØļ¼╝ņØś ņāØļ¼╝ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ļīĆņ╣śļ░░ņ¢æņØä ĒåĄĒĢ┤ ņäĀļ░£ļÉ£ ĻĖĖĒĢŁĻĘĀņØä ļīĆņāüņ£╝ļĪ£ ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æņŚÉ ļīĆĒĢ£ ņāØļ¼╝ĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢ£ Ļ▓░Ļ│╝, DF30, DF137, DM48, DH2, DH16 ĻĘĀņŻ╝ļŖö 80.4% ņØ┤ņāüņØś ļåÆņØĆ ļ░®ņĀ£Ļ░Ćļź╝ ļ│┤ņśĆņ£╝ļ®░, ļ»ĖņāØļ¼╝ņĀ£ĒÆł EXTN-1 (B. vallismortis EXTN-1)Ļ│╝ ĒÖöĒĢÖļåŹņĢĮ fludioxonilņØĆ Ļ░üĻ░ü 35.1%, 95.8%ņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. ļ░░ņČö ļ┐īļ”¼Ēś╣ļ│æņŚÉ ļīĆĒĢ£ ĻĖĖĒĢŁĻĘĀņØś ņāØļ¼╝ĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢ£ Ļ▓░Ļ│╝, DF30, DM142, DM146, DM152, DH2 ĻĘĀņŻ╝ļŖö 83.0% ņØ┤ņāüņØś ļåÆņØĆ ļ░®ņĀ£Ļ░Ćļź╝ ļ│┤ņśĆņ£╝ļ®░, ļ»ĖņāØļ¼╝ņĀ£ĒÆł EXTN-1Ļ│╝ ĒÖöĒĢÖļåŹņĢĮ cyazopamidļŖö Ļ░üĻ░ü 73.5%, 99.3%ņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. Ļ│ĀņČö ĒāäņĀĆļ│æņŚÉ ļīĆĒĢ£ ĻĖĖĒĢŁĻĘĀņØś ņāØļ¼╝ĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢ£ Ļ▓░Ļ│╝, DF30, DM142, DM146, DM152 ĻĘĀņŻ╝ļŖö 82.3% ņØ┤ņāüņØś ļåÆņØĆ ļ░®ņĀ£Ļ░Ćļź╝ ļ│┤ņśĆņ£╝ļ®░, ļ»ĖņāØļ¼╝ņĀ£ĒÆł EXTN-1Ļ│╝ ĒÖöĒĢÖļåŹņĢĮ propinebļŖö Ļ░üĻ░ü 86.6%, 47.2%ņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. Ļ│ĀņČö ņŚŁļ│æņŚÉ ļīĆĒĢ£ ĻĖĖĒĢŁĻĘĀņØś ņāØļ¼╝ĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢ£ Ļ▓░Ļ│╝, DM152, DH16ņØś ĻĘĀņŻ╝ļŖö 83.0% ņØ┤ņāüņØś ļ░®ņĀ£Ļ░Ćļź╝ ļ│┤ņśĆņ£╝ļ®░, ļ»ĖņāØļ¼╝ņĀ£ĒÆł EXTN-1Ļ│╝ ĒÖöĒĢÖļåŹņĢĮ dimetomorphņØĆ Ļ░üĻ░ü 65.5%, 98.9%ņØś ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. ļö░ļØ╝ņä£, B. cinerea, P. brassicae, C. acutatum, P. capsiciņŚÉ ļīĆĒĢśņŚ¼ ņāØļ¼╝ĒÖ£ņä▒ņØ┤ ņÜ░ņłśĒĢ£ 10ņóģņØś ĻĖĖĒĢŁĻĘĀņØä ņäĀļ░£ĒĢśņśĆĻ│Ā, ņāØļ¼╝ņĀü ļ░®ņĀ£ņĀ£ļĪ£ ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż(Table 2).
ņØ┤ ļ░¢ņØś, BacillusņåŹ, PaenibacillusņåŹ, StreptomycesņåŹ ĻĘĀņŻ╝ļŖö ļŗżņ¢æĒĢ£ ĻĖ░ņŻ╝ņÖĆ ņŗØļ¼╝ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢśņŚ¼ ņāØļ¼╝ņĀü ļ░®ņĀ£ ĒÜ©Ļ│╝Ļ░Ć ļ│┤Ļ│ĀļÉśĻ│Ā ņ׳ļŗż. XuņÖĆ Kim (2014)ņØĆ ĒåĀņ¢æņŚÉņä£ ļČäļ”¼ĒĢ£ P. polymyxa SC09-21ņØĆ ĒåĀļ¦łĒåĀ ĻĘ╝ļČĆņŹ®ņØīļ│æņŚÉ ļīĆĒĢśņŚ¼ ņé┤ĻĘĀņĀ£ metconazoleĻ│╝ ļ╣äĻĄÉĒĢśņŚ¼ 81.7%ņØś ņÜ░ņłśĒĢ£ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļāłĻ│Ā, Chen ļō▒ (2000)ņØĆ ņäĀņČ®ņØä ņ¢ĄņĀ£ĒĢśļŖö ĒåĀņ¢æņŚÉņä£ ļČäļ”¼ļÉ£ S. costaricanus ĻĘĀņŻ╝ļŖö ĻĖĖĒĢŁņĀü ņ×æņÜ®ņØś ĒŖ╣ņä▒ņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ņ¢┤ ņäĀņČ®ņŚÉ ļīĆĒĢ£ ņāØļ¼╝ņĀü ļ░®ņĀ£ņĀ£ļĪ£ Ļ▓Ćņ”ØĒĢśņśĆļŗż. ļśÉĒĢ£, S. misionensisņØś ļæÉ ĻĘĀņŻ╝ BH4-1, BH4-3ņØĆ in vitroņÖĆ in vivoņŚÉņä£ Ēö╝ņŖżĒāĆņ╣śņśż ņżäĻĖ░ļ¦łļ”äļ│æ, ĻČżņ¢æ ļ│æņŚÉ ļīĆĒĢśņŚ¼ ņé┤ĻĘĀņĀ£ Cupravit (Bayer CropScience, Granada, Mexico)Ļ│╝ ļ╣äĻĄÉĒĢśņŚ¼ ļ╣äļō▒ĒĢ£ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļ│┤ņŚ¼ ņāØļ¼╝ņĀü ļ░®ņĀ£ļĀźņØä Ļ▓Ćņ”ØĒĢśņśĆļŗż(Torabi ļō▒, 2019). ĒŖ╣Ē׳, BacillusņåŹ ĻĘĀņØĆ ņāØļ¼╝ņĀü ļ░®ņĀ£ņŚÉ ņżæņÜöĒĢ£ ņŚŁĒÖ£ņØä ĒĢśļŖö iturin, surfactin, bacillomycin, fengycin ļō▒ņØś lipopeptideĻ│äļź╝ ĒżĒĢ©ĒĢ£ ĒĢŁņāØļ¼╝ņ¦łņØä ļČäļ╣äĒĢ£ļŗżĻ│Ā ņĢīļĀżņĀĖ ņ׳ļŗż(Ntushelo ļō▒, 2019; Zeriouh ļō▒, 2011). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ļÅä ņäĀļ░£ļÉ£ 4ņóģņØś BacillusņåŹ ĻĘĀņØĆ iturin AņÖĆ surfactinĻ│╝ Ļ░ÖņØĆ ĒĢŁņāØļ¼╝ņ¦łņØä ņāØņé░ĒĢĀ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż.
ĻĖĖĒĢŁļ»ĖņāØļ¼╝ņØś ļÅÖņĀĢ
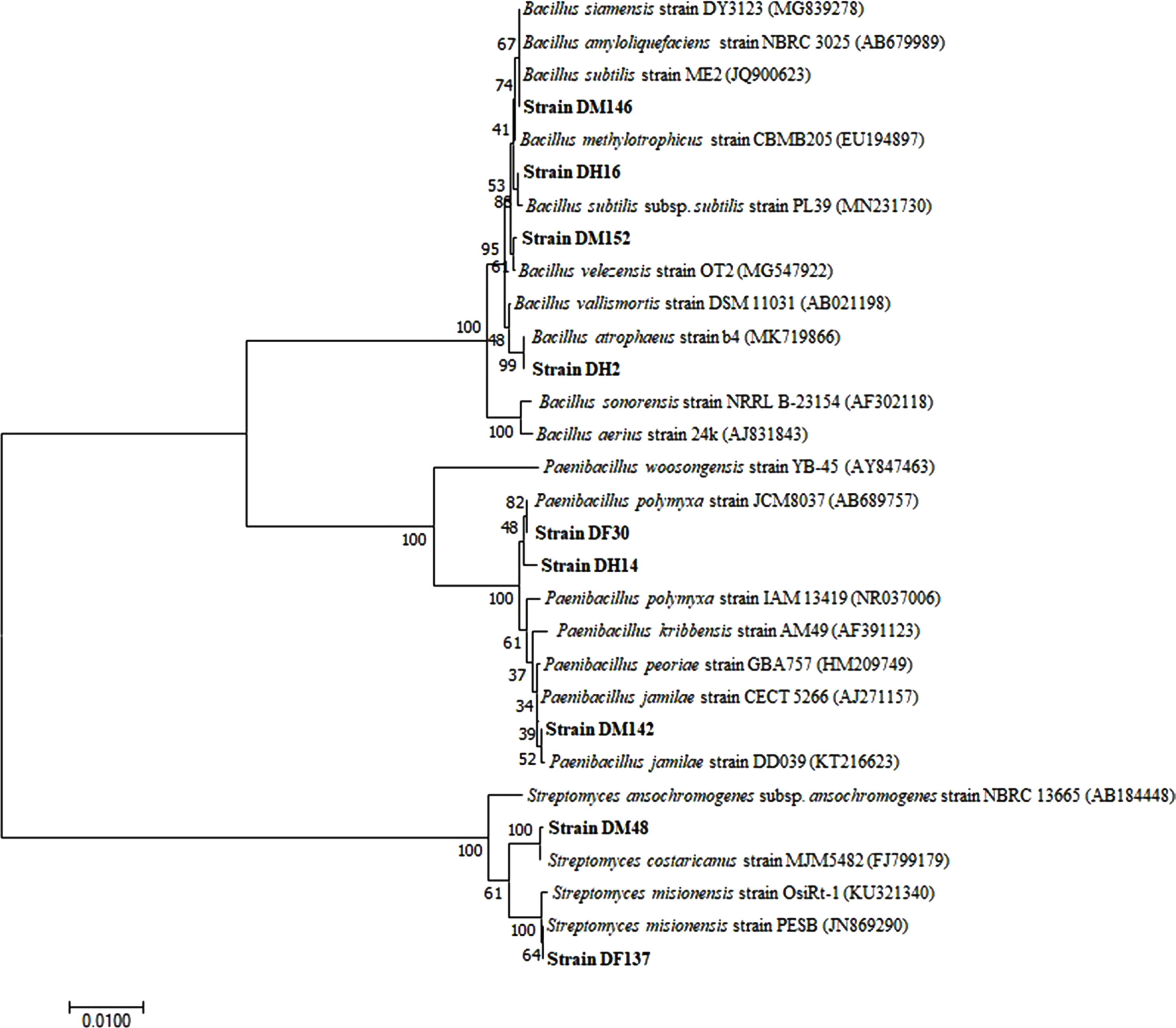
ņäĀļ░£ļÉ£ 10ņóģņØś ĻĖĖĒĢŁĻĘĀņØĆ ņŻ╝ņé¼ņĀäņ×ÉĒśäļ»ĖĻ▓ĮņØä ĒåĄĒĢ£ ĒśĢĒā£ĒĢÖņĀü ļČäņäØĻ│╝ 16s rRNA ņŚ╝ĻĖ░ņä£ņŚ┤ ļČäņäØņØä ĒåĄĒĢśņŚ¼ ļÅÖņĀĢĒĢ£ Ļ▓░Ļ│╝, 4ņóģņØś Bacillus spp., 3ņóģņØś Paenibacillus spp., 3ņóģņØś Streptomyces spp.ņ£╝ļĪ£ ļČäļźśļÉśņŚłļŗż. DM146 ĻĘĀņŻ╝ļŖö B. subtilis, DM152 ĻĘĀņŻ╝ļŖö B. velezensis, DH16 ĻĘĀņŻ╝ļŖö B. subtilis subsp. subtilisņÖĆ 99.93% ņāüļÅÖņä▒ņØä ļ│┤ņśĆĻ│Ā, DH2 ĻĘĀņŻ╝ļŖö B. atrophaeusņÖĆ 100% ņāüļÅÖņä▒ņØä ļ│┤ņśĆļŗż. ļśÉĒĢ£, DF30, DH14 ĻĘĀņŻ╝ļŖö P. polymyxaņÖĆ 99.86% ņāüļÅÖņä▒ņØ┤ ņØ╝ņ╣śĒĢśņśĆĻ│Ā, DM142 ĻĘĀņŻ╝ļŖö P. jamilaeņÖĆ 100% ņāüļÅÖņä▒ņØ┤ ņØ╝ņ╣śĒĢśņśĆļŗż. ļśÉĒĢ£, DF137 ĻĘĀņŻ╝ļŖö S. misionensisņÖĆ 99.85% ņāüļÅÖņä▒ņØä ļ│┤ņśĆĻ│Ā, DM48 ĻĘĀņŻ╝ļŖö S. costaricanusņÖĆ 99.93% ņāüļÅÖņä▒ņØ┤ ņØ╝ņ╣śĒĢśņśĆļŗż. DH92 ĻĘĀņŻ╝ļŖö genomic DNAļź╝ ĒÜŹļōØĒĢśļŖöļŹ░ ņŗżĒī©ĒĢśņŚ¼ 16S rRNA gene ņŚ╝ĻĖ░ņä£ņŚ┤ņØä ļČäņäØĒĢśņ¦Ć ļ¬╗ĒĢśņśĆņ¦Ćļ¦ī, ĒśĢĒā£ņĀü ĒŖ╣ņä▒ņŚÉ ļö░ļØ╝ Streptomyces sp.ļĪ£ ļÅÖņĀĢĒĢśņśĆļŗż(Figs. 1, 2).
BacillusņåŹ, PaenibacillusņåŹ, StreptomycesņåŹ ĻĘĀņŻ╝ļŖö ļŗżņ¢æĒĢ£ ņŗØļ¼╝ ĻĘĀļźśļ│æņŚÉ ļīĆĒĢśņŚ¼ Ēżņ×É ļ░£ņĢäņÖĆ ĻĘĀņé¼ ņāØņןņØä ņ¢ĄņĀ£ĒĢśņŚ¼ Ļ░ĢĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä Ļ░Ćņ¦ĆĻ│Ā ņ׳ņ¢┤ ņāØļ¼╝ņĀü ļ░®ņĀ£ļŖźņØ┤ ņÜ░ņłśĒĢ£ Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ļŗż(Huo ļō▒, 2010; Lee ļō▒, 2006; Li ļō▒, 2012). ļśÉĒĢ£, ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ļČäļ”¼ļÉ£ ņןļé┤ļ»ĖņāØļ¼╝ņØĆ BacillusņåŹ, PaenibacillusņåŹ ĻĘĀņŻ╝Ļ░Ć ņĀ£ņØ╝ ļ¦ÄņØĆ ļČäļ”¼ļ╣äļź╝ ļ│┤ņśĆļŗż(data not shown). ņØ┤ļ¤¼ĒĢ£ BacillusņåŹ, PaenibacillusņåŹ ĻĘĀņŻ╝ļōżņØĆ endosporeļź╝ ĒśĢņä▒ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ Ļ│Āņś©, ļ░®ņé¼ņäĀ, Ļ▒┤ņĪ░ ļō▒ ļČłļ”¼ĒĢ£ ĒÖśĻ▓ĮņŚÉņä£ ņāØņןņØä Ļ░ĆļŖźĒĢśĻ▓ī ĒĢ£ļŗż(McSpadden Gardener, 2004; Nicholson ļō▒, 2000). ņØ┤ņÖĆ ļ¦łņ░¼Ļ░Ćņ¦ĆļĪ£ Ļ│żņČ®ņØĆ ņäŁņŗØ ĒÖśĻ▓ĮņŚÉ ļö░ļØ╝ ņĀĢņ░®ĒĢśĻ│Ā ņ׳ļŖö ņןļé┤ ļ»ĖņāØļ¼╝ ĻĄ░ņØś ņ¢æņāüņØ┤ ļŗżļź┤ļ®░, ņØ┤ļ¤¼ĒĢ£ Ļ│ĄņāØņ×ÉļŖö Ļ│Ąņ¦äĒÖöļź╝ ĒåĄĒĢśņŚ¼ ņśüņ¢æņåīĻ░Ć ļČłļ”¼ĒĢ£ ĻĘ╣ĒĢ£ ĒÖśĻ▓ĮņŚÉņä£ļÅä ņä£ņŗØ Ļ░ĆļŖźĒĢśĻ▓ī ĒĢ£ļŗż. ļö░ļØ╝ņä£, Ļ│żņČ®ņØś ņןļé┤ ļ»ĖņāØļ¼╝ ņżæ endosporeļź╝ ĒśĢņä▒ĒĢśļŖö ĻĘĀņŻ╝ļōżņØĆ ņŗØļ¼╝ ļ│æņøÉĻĘĀņØä ņ¢ĄņĀ£ĒĢśĻ▒░ļéś ņ×æļ¼╝ņØś ņāØņ£ĪņØä ņ¦üŌĆóĻ░äņĀæņĀüņ£╝ļĪ£ ņ┤ēņ¦äņŗ£ĒéżļŖö ļō▒ ņ£ĀņÜ®ĒĢ£ ĒÜ©Ļ│╝ļź╝ ļåŹņŚģ ĒÖśĻ▓ĮņŚÉ ĻĖ░ņŚ¼ĒĢĀ ņłś ņ׳ļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż. ļśÉĒĢ£, Han ļō▒(2015)ņØĆ Ļ░ÖņØĆ BacillusņåŹņØ╝ņ¦ĆļØ╝ļÅä strainņŚÉ ļö░ļØ╝ Ļ░üĻ░ü ļČäļ╣äĒĢśļŖö ĒĢŁņāØļ¼╝ņ¦łņØ┤ ļŗżļź┤Ļ│Ā ĒĢŁĻĘĀĒÖ£ņä▒ņØś ĒÜ©Ļ│╝Ļ░Ć ļŗżļź╝ ņłś ņ׳ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż. ņØ┤ņÖĆ Ļ░ÖņØ┤ ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś Ļ░üĻ░üņØś ņåīĒÖöĻĖ░Ļ┤Ć ņĀäņן, ņżæņן, ĒøäņןņŚÉņä£ strainņØ┤ Ļ░ÖļŹöļØ╝ļÅä ņןļé┤ ĒŖ╣ņä▒ņŚÉ ļö░ļØ╝ ņĀĢņ░® ļ»Ė ņāØļ¼╝ņØś ĻĖ░ļŖźņä▒ņØ┤ ļŗżļź╝ Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż.
IAA ņāØņä▒ļŖź
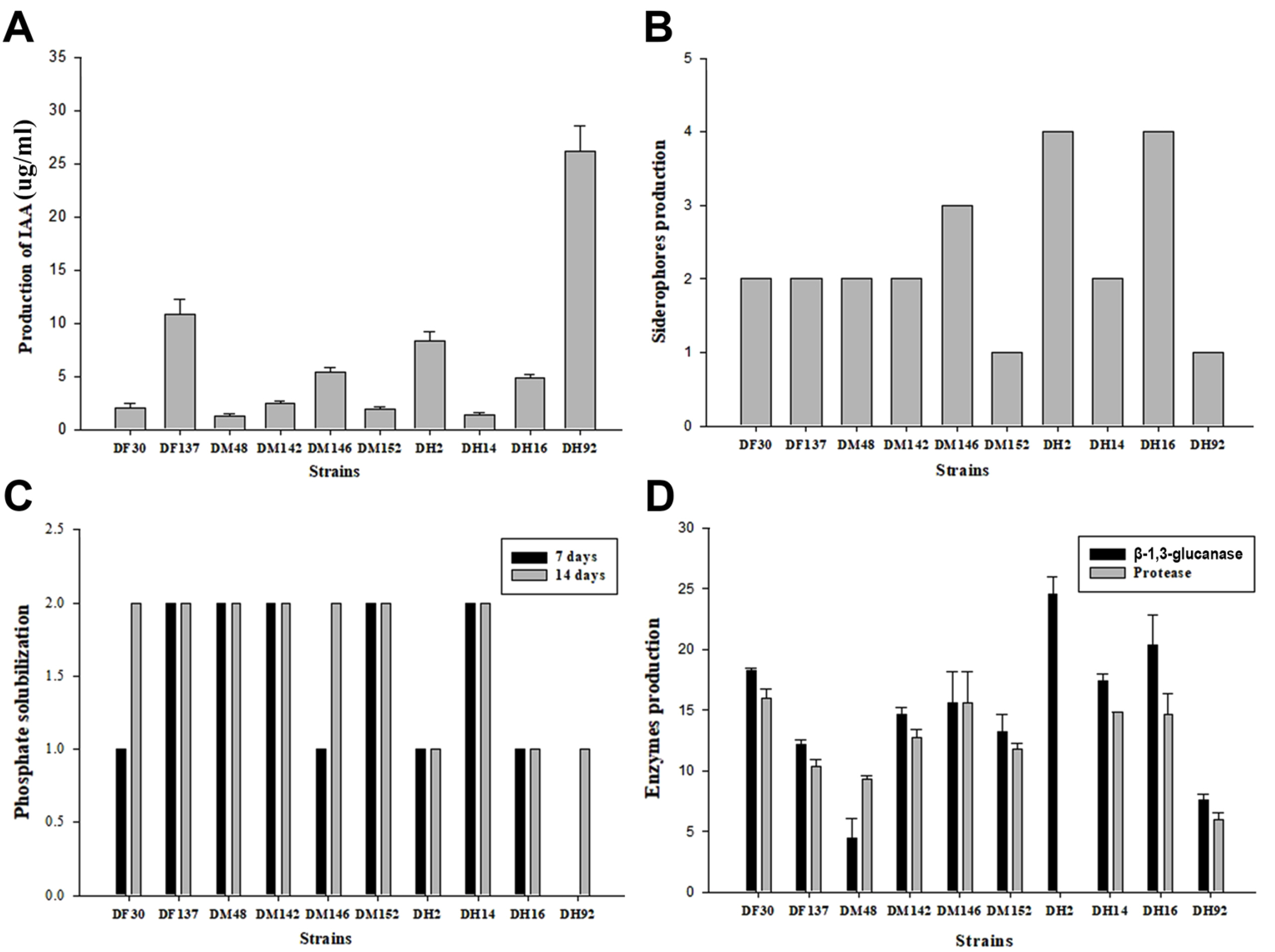
ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņŚÉ ņØśĒĢ£ ņŗØļ¼╝ ĒśĖļź┤ļ¬¼ņØś ņāØņé░ņØĆ ņŗØļ¼╝ņāØņןņ┤ēņ¦ä ĒÖ£ņä▒ņØś Ļ░Ćņן ņżæņÜöĒĢ£ ņÜöņåī ņżæ ĒĢśļéśņØ┤ļŗż. ņ¦ĆĻĖłĻ╣īņ¦Ć IAA, cytokinins, gibberellins Ļ░ÖņØĆ ņŗØļ¼╝ĒśĖļź┤ļ¬¼ņØä ņāØņé░ĒĢśļŖö ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØś ĒÜ©Ļ│╝ņŚÉ ļīĆĒĢ£ ļ¦ÄņØĆ ļ│┤Ļ│ĀĻ░Ć ņ׳ļŗż(Kavamura ļō▒, 2013; Son ļō▒, 2014). ņäĀļ░£ļÉ£ 10ņóģņØś ĻĘĀņŻ╝ņŚÉ ļīĆĒĢ£ IAA ņāØņä▒ ĒÖ£ņä▒ ņĪ░ņé¼ļź╝ ņ£äĒĢ┤ ĒØĪĻ┤æļÅäļź╝ ņĖĪņĀĢĒĢ£ Ļ▓░Ļ│╝, 1.2ŌĆÆ26.9 ╬╝g/mlņØś ņÜ░ņłśĒĢ£ IAA ņāØņä▒ļŖźņØä ļ│┤ņśĆļŗż. ĒøäņןņŚÉņä£ ļČäļ”¼ĒĢ£ DH92 ĻĘĀņŻ╝Ļ░Ć 26.9 ╬╝g/mlļĪ£ Ļ░Ćņן ļåÆņØĆ ņāØņä▒ļŖźņØä ļ│┤ņśĆĻ│Ā, ņżæņןņŚÉņä£ ļČäļ”¼ĒĢ£ DM48 ĻĘĀņŻ╝Ļ░Ć 1.2 ╬╝g/mlļĪ£ Ļ░Ćņן ļé«ņØĆ ņāØņä▒ļŖźņØä ļ│┤ņśĆļŗż(Fig. 3A). IAA ņāØņä▒ņØĆ ņäĖĒż Ēś╣ņØĆ ņŗØļ¼╝ņØś ņāØņןĻ│╝ņĀĢņØä ņĀ£ņ¢┤ĒĢśĻĖ░ ļĢīļ¼ĖņŚÉ ņżæņÜöĒĢ£ ĒŖ╣ņ¦ĢņØ┤ļŗż(Lambrecht ļō▒, 2000). ņØ┤ļ¤¼ĒĢ£ Ļ▓░Ļ│╝ļŖö ļŗżņ¢æĒĢ£ ĻĖ░ļŖźņä▒ņØä ļ│┤ņ£ĀĒĢ£ ņןļé┤ ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØ┤ ņŗØļ¼╝Ļ│╝ ļ»ĖņāØļ¼╝ņØś ņāüĒśĖņ×æņÜ®ņØä ĒåĄĒĢśņŚ¼ ņŗØļ¼╝ņØś ņāØņןņØä ņ┤ēņ¦äĒĢ£ļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż.
ņØĖņé░ Ļ░ĆņÜ®ĒÖö ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņØĖņé░ņØĆ ņŚÉļäłņ¦Ć ļīĆņé¼ņŚÉ Ļ┤ĆņŚ¼ĒĢ©ņ£╝ļĪ£ņŹ© ņŗØļ¼╝ņØś ņāØņןĻ│╝ ļ░£ļŗ¼ņŚÉ Ļ░Ćņן ĒĢäņłśņĀüņØĖ ņÜöņåī ņżæ ĒĢśļéśņØ┤ļŗż. ņØ┤ļ¤¼ĒĢ£ ņØĖņé░ņØĆ ĒåĀņ¢æņŚÉ ļ¼┤ĻĖ░ ņØĖņé░ņŚ╝ ĒśĢĒā£ļĪ£ ņĪ┤ņ×¼ĒĢśņ¦Ćļ¦ī ļČłņÜ®ĒÖöļÉśņ¢┤ ņŗØļ¼╝ņØ┤ ņ¦üņĀæņĀüņ£╝ļĪ£ ĒØĪņłśļź╝ ĒĢĀ ņłś ņŚåļŗż. ļö░ļØ╝ņä£ ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØś ņĀüņÜ®ņØĆ ļé£ņÜ®ņä▒ ņØĖņé░ņŚ╝ņØä ļČäĒĢ┤ĒĢśņŚ¼ ņŗØļ¼╝ņØ┤ ĒØĪņłśĒĢĀ ņłś ņ׳Ļ▓ī ļÅäņøĆņØä ņżä ņłś ņ׳ņ£╝ļ®░, ĒÖöĒĢÖļ╣äļŻīņØś ņé¼ņÜ®ņØä ņżäņØ┤Ļ│Ā ĒåĀņ¢æ ļČłĻĘĀĒśĢņØä ļ¦ēņØä ņłś ņ׳ļŗż(Hameeda ļō▒, 2008; Rajan ļō▒, 1996). ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņäĀļ░£ļÉ£ 10ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ ļ¬©ļæÉ ļČłņÜ®ņä▒ ņØĖņé░ņØ┤ ĒżĒĢ©ļÉ£ PikovskayaŌĆÖs agar ļ░░ņ¦ĆņŚÉņä£ Ēł¼ļ¬ģĒÖśņØä ĒśĢņä▒ĒĢśņśĆļŗż. DF137, DM48, DM152, DH14 ĻĘĀņŻ╝ļŖö 7ņØ╝, 14ņØ╝ Ēøä ļ¬©ļæÉ ļåÆņØĆ ĒÖ£ņä▒ņØä ļ│┤ņśĆņ£╝ļ®░, DH2, DH92 ĻĘĀņŻ╝ļŖö 14ņØ╝ ĒøäņŚÉļÅä ļé«ņØĆ ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż(Fig. 3B).
Siderophore ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņ▓ĀņØĆ ĒĢäņłśņĀüņØĖ ņśüņ¢æņåīņØ┤ņ¦Ćļ¦ī ĒåĀņ¢æņŚÉņä£ ņāüļīĆņĀüņ£╝ļĪ£ ņēĮĻ▓ī ņÜ®ĒĢ┤ļÉśņ¦Ć ņĢŖļŖöļŗż(Vessey, 2003). ņØ┤ļŖö ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņŚÉ ņØśĒĢ┤ ņĀ£ĒĢ£ļÉ£ ņ▓Ā(Fe3+) ņä▒ļČä Ļ▓░ĒĢ®ļ¼╝ņ¦łņØĖ siderophoreļź╝ ņāØņé░ĒĢśņŚ¼ ļŗżņ¢æĒĢ£ ņŗØļ¼╝ļ│æņøÉĻĘĀņØś ņāØņ£ĪņØä ņĀĆĒĢ┤ĒĢśĻ▒░ļéś ņ¦üĻ░äņĀæņĀüņ£╝ļĪ£ ņ×æļ¼╝ņØś ņāØņןņØä ņ┤ēņ¦äĒĢĀ ņłś ņ׳ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņäĀļ░£ļÉ£ 10ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ ļ¬©ļæÉ CAS agar ļ░░ņ¦ĆņŚÉņä£ ņśżļĀīņ¦Ć ņāēĻ╣öļĪ£ ļ│ĆņāēļÉśņ¢┤ halo zoneņØä ĒśĢņä▒ĒĢśņśĆļŗż. DM146, DH2, DH16 ĻĘĀņŻ╝Ļ░Ć CAS agar ļ░░ņ¦ĆņŚÉņä£ 15.1 mm ņØ┤ņāüņØś halo zoneņØä ĒśĢņä▒ĒĢśņŚ¼ siderophore ņāØņä▒ļŖźņØ┤ ņÜ░ņłśĒĢ£ Ļ▓āņ£╝ļĪ£ ĒÅēĻ░ĆļÉśņŚłļŗż. DM152, DH92 ĻĘĀņŻ╝ļŖö 0.1ŌĆÆ5 mmņØś halo zoneņØä ĒśĢņä▒ĒĢśņŚ¼ ļé«ņØä ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż(Fig. 3C). ņØ┤ļŖö ņ£ĀņÜ® ņןļé┤ ļ»ĖņāØļ¼╝ņŚÉ ņØśĒĢ£ siderophoreņØś ņāØņé░ņØĆ ņŗØļ¼╝ņŚÉ ņ▓ĀņØä Ļ│ĄĻĖēĒĢĀ ņłś ņ׳ļŖö ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢĀ ņłś ņ׳ļŗżĻ│Ā ņāØĻ░üļÉ£ļŗż.
Ļ░ĆņłśļČäĒĢ┤ĒÜ©ņåī(╬▓-1,3-glucanase, protease) ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņāØļ¼╝ņĀü ļ░®ņĀ£ļŖźĻ│╝ ņŗØļ¼╝ņāØņןņ┤ēņ¦äļŖźņØä Ļ░Ćņ¦ĆļŖö ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØś ĒŖ╣ņ¦Ģ ņżæ ĒĢśļéśļŖö ņŗØļ¼╝ļ│æņøÉĻĘĀņ£╝ļĪ£ļČĆĒä░ ņŗØļ¼╝ņØä ļ│┤ĒśĖĒĢĀ ņłś ņ׳ļŖö ╬▓ -1,3-glucanase, protease Ļ░ÖņØĆ ņ¦äĻĘĀ ņÖĖļ¦ē Ļ░ĆņłśļČäĒĢ┤ ĒÜ©ņåīņØś ĒÖ£ņä▒ņØ┤ļŗż. ļ│Ė ņŚ░ĻĄ¼ņŚÉņä£ ņäĀļ░£ļÉ£ 10ņóģņØś ņ£ĀņÜ® ņןļé┤ ļ»ĖņāØļ¼╝ņØĆ ļŗżņ¢æĒĢ£ ņłśņżĆņØś ╬▓-1,3-glucanase, protease ĒÖ£ņä▒ ĒÜ©Ļ│╝ļź╝ ļ│┤ņśĆļŗż. DF30, DF137, DM142, DM146, DM152, DH14, DH16 ĻĘĀņŻ╝ļŖö 10.0 mm ņØ┤ņāüņØś Ēł¼ļ¬ģĒÖśņØä ĒśĢņä▒ĒĢśņŚ¼ ļåÆņØĆ ĒÖ£ņä▒ņØä ļéśĒāĆļāłļŗż. DH2 ĻĘĀņŻ╝ļŖö ╬▓-1,3-glucanase ĒÜ©ņåī ĒÖ£ņä▒ņØĆ ņÜ░ņłśĒĢśņśĆņ¦Ćļ¦ī, protease ĒÖ£ņä▒ņØĆ ļéśĒāĆļé┤ņ¦Ć ņĢŖņĢśļŗż(Fig. 3D). ╬▓-1,3-glucanase, protease ĒÖ£ņä▒ ĒÜ©Ļ│╝ļź╝ Ļ░Ćņ¦ä ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØĆ ņŗØļ¼╝ņāØņן ņ┤ēņ¦äĻ│╝ ņ£ĀĻĖ░ļ¼╝ ļČäĒĢ┤ņŚÉ ņ׳ņ¢┤ ņżæņÜöĒĢ£ ņŚŁĒĢĀņØä ĒĢĀ ļ┐Éļ¦ī ņĢäļŗłļØ╝ ņŗØļ¼╝ļ│æņøÉĻĘĀņØś ņāØņןņØä ņ¢ĄņĀ£ĒĢśļŖö ļŹ░ ļÅäņøĆņØä ņżä ņłś ņ׳ļŗż(Minaxi ļō▒, 2012).
ņŗØļ¼╝ņāØņןņ┤ēņ¦ä ĒÖ£ņä▒ Ļ▓ĆņĀĢ
ņäĀļ░£ļÉ£ 10ņóģņØś ĻĘĀņŻ╝ļź╝ ĒåĀļ¦łĒåĀņÖĆ Ļ│ĀņČö ņ£Āļ¼śļź╝ ļīĆņāüņ£╝ļĪ£ ņŗØļ¼╝ņāØņןņ┤ēņ¦ä ĒÖ£ņä▒ņØä Ļ▓ĆņĀĢĒĢśņśĆļŗż. ĒåĀļ¦łĒåĀ ņ£Āļ¼śņŚÉ ņ▓śļ”¼ĒĢ£ ĻĖĖĒĢŁĻĘĀ ņżæ DF30, DM152, DH16 ĻĘĀņŻ╝Ļ░Ć ĒåĀļ¦łĒåĀ ņŗØļ¼╝ņ▓┤ņØś ļ¬©ļōĀ ĻĖ░Ļ┤ĆņŚÉņä£ ņāØņןņ┤ēņ¦äĒÖ£ņä▒ņØ┤ Ļ░Ćņן ņÜ░ņłśĒĢśĻ▓ī ļéśĒāĆļāłļŗż. ņżäĻĖ░ ĻĖ░Ļ┤ĆņØś ĻĖĖņØ┤ņÖĆ ņ¦üĻ▓ĮņØĆ DF30 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć 54.32 cm, 7.30 mmļĪ£ Ļ░Ćņן ļåÆņĢśņ£╝ļ®░, ņāØņ▓┤ ņżæļ¤ēĻ│╝ Ļ▒┤ņĪ░ ņżæļ¤ēņŚÉņä£ļŖö DM152 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć 56.56 g, 3.22 gņ£╝ļĪ£ Ļ░Ćņן ļåÆņĢśļŗż. ļ┐īļ”¼ ĻĖ░Ļ┤ĆņØś Ļ▓ĮņÜ░ ĻĖĖņØ┤ļŖö DF137 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć Ļ░Ćņן ļåÆņĢśņ£╝ļ®░, ņāØņ▓┤ ņżæļ¤ēĻ│╝ Ļ▒┤ņĪ░ ņżæļ¤ēņŚÉņä£ļŖö DM152 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć 8.84 g, 0.55 gņ£╝ļĪ£ Ļ░Ćņן ņóŗņØĆ ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż. ņŚĮļĪØņåī ĒĢ©ļ¤ēņŚÉņä£ļŖö DM142 ĻĘĀņŻ╝ ņ▓śļ”¼ĻĄ¼Ļ░Ć 37.63 spad unitsļĪ£ Ļ░Ćņן ņóŗņØĆ ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż(Table 3). Ļ│ĀņČö ņ£Āļ¼śņØś Ļ▓ĮņÜ░, ņ▓śļ”¼ĻĄ¼ ņżæņŚÉņä£ DM142, DM152Ļ░Ć ņ▓śļ”¼ļÉ£ Ļ│ĀņČöņØś ļ¬©ļōĀ ĻĖ░Ļ┤ĆņŚÉņä£ ņāØņןņ┤ēņ¦äĒÖ£ņä▒ņØ┤ Ļ░Ćņן ņÜ░ņłśĒĢśĻ▓ī ļéśĒāĆļāłļŗż. ņżäĻĖ░ņØś ĻĖĖņØ┤ņÖĆ ņżæļ¤ēņŚÉņä£ļŖö DM142 ĻĘĀņŻ╝Ļ░Ć 32.78 cm, 6.90 mmļĪ£ Ļ░Ćņן ļåÆņĢśņ£╝ļ®░, ņāØņ▓┤ ņżæļ¤ēĻ│╝ Ļ▒┤ņĪ░ ņżæļ¤ēņŚÉņä£ļŖö DM152 ĻĘĀņŻ╝Ļ░Ć 22.96 g, 2.91 g, DM142 ĻĘĀņŻ╝Ļ░Ć 21.62 g, 3.37 gņ£╝ļĪ£ Ļ░Ćņן ļåÆņĢśļŗż. ļ┐īļ”¼ ĻĖ░Ļ┤ĆņØś Ļ▓ĮņÜ░ ĻĖĖņØ┤ļŖö DM152 ĻĘĀņŻ╝Ļ░Ć 19.2 cmļĪ£ Ļ░Ćņן ļåÆņĢśņ£╝ļ®░, ņāØņ▓┤ ņżæļ¤ēĻ│╝ Ļ▒┤ņĪ░ ņżæļ¤ēņŚÉņä£ļŖö DM142 ĻĘĀņŻ╝Ļ░Ć 13.62 g, 1.47 gņ£╝ļĪ£ Ļ░Ćņן ļåÆņĢśļŗż. ņŚĮļĪØņåī ĒĢ©ļ¤ēņŚÉņä£ļŖö ņ£ĀņØśņ░©ņØ┤ļŖö ņŚåņŚłņ¦Ćļ¦ī DM152 ĻĘĀņŻ╝Ļ░Ć 56.22 spad unitsļĪ£ Ļ░Ćņן ņóŗņØĆ ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż(Table 4). ņäĀļ░£ļÉ£ 10ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ ņżæņŚÉņä£, DM152 ĻĘĀņŻ╝ļŖö ņØĖņé░ Ļ░ĆņÜ®ĒÖö ĒÖ£ņä▒ņØĆ ņÜ░ņłśĒĢśņ¦Ćļ¦ī IAA, siderophore ņāØņä▒ ĒÖ£ņä▒ņØ┤ ļŗżļźĖ ĻĘĀņŻ╝ņÖĆ ļ╣äĻĄÉĒĢśņŚ¼ ņĀĆņĪ░ĒĢ©ņŚÉļÅä ļČłĻĄ¼ĒĢśĻ│Ā Ļ│ĀņČöņÖĆ ĒåĀļ¦łĒåĀ ņŗØļ¼╝ņ▓┤ ļ¬©ļæÉņŚÉņä£ Ļ░Ćņן Ļ░ĢļĀźĒĢ£ ņŗØļ¼╝ ņāØņן ņ┤ēņ¦ä ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż. ņØ┤ļŖö ļŗżņ¢æĒĢ£ ļ®öņ╗żļŗłņ”śņØ┤ ņāüĒśĖņ×æņÜ®ĒĢśĻ▒░ļéś ĒĢśļéśņØś ļÅģļ”ĮļÉ£ ļ®öņ╗żļŗłņ”śņØ┤ ņŗØļ¼╝ ņāØņן ņ┤ēņ¦äĒĢśļŖö ĒÜ©Ļ│╝ņŚÉ Ļ┤ĆņŚ¼Ē¢łņØä Ļ▓āņ£╝ļĪ£ ņČöņĀĢļÉ£ļŗż. ņØ┤ļŖö Son ļō▒(2014)ņØ┤ ļ│┤Ļ│ĀĒĢ£ Ļ│ĀņČö ņŗØļ¼╝ņ▓┤ņŚÉ ņ▓śļ”¼ĒĢ£ Kluyvera cryocrescens KUDC1771 ĻĘĀņŻ╝ņÖĆ ņØ╝ņ╣śĒĢ£ļŗż. ņØ┤ļŖö ņŗØļ¼╝ ņāØņן ņ┤ēņ¦äņŚÉ Ļ┤ĆņŚ¼ĒĢśļŖö ļ®öņ╗żļŗłņ”śņØä ĻĘ£ļ¬ģĒĢśĻĖ░ ņ£äĒĢ┤ ļéśņĢäĻ░Ć ļŗżņ¢æĒĢ£ ņŚ░ĻĄ¼Ļ░Ć ĒĢäņÜöĒĢśļŗż.
Ļ│żņČ®ņØĆ ņןļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö ļ»ĖņāØļ¼╝Ļ│╝ ņāüĒśĖņ×æņÜ®ņØä ĒĢśļ®┤ņä£ ņ£ĀņØĄĒĢ£ Ļ│ĄņāØĻ┤ĆĻ│äļź╝ ņ£Āņ¦ĆĒĢśļ®░, ņןļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö Ļ│ĄņāØņ×ÉļŖö Ļ│żņČ®ņŚÉ ņ×Āņ×¼ņĀüņØĖ ņŚŁĒĢĀļĪ£ ņāØņ£ĪņŚÉ ĒĢäņłśņĀüņØĖ ņśüĒ¢źņØä ļ»Ėņ╣śļ®░ ĻĘ╣ĒĢ£ ĒÖśĻ▓ĮņŚÉļÅä ņĀüņØæĒĢĀ ņłś ņ׳ļÅäļĪØ ļÅäņÖĆņżĆļŗż. ņĄ£ĻĘ╝ ņØ┤ļ¤¼ĒĢ£ ņØ┤ņ£ĀļĪ£ ņןļé┤ ļ»ĖņāØļ¼╝ņØä ņ£ĀņÜ®ĒĢ£ ņåīņ×¼ļĪ£ ņØĖņŗØĒĢśĻ│Ā ņ׳ņ£╝ļ®░, ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņĀ£ņŚÉ ļīĆĒĢ£ Ļ▓Ćņ”ØņØ┤ ļ¦ÄņØĆ ļģ╝ļ¼ĖņŚÉņä£ ļ│┤Ļ│ĀļÉśņŚłļŗż. Lee ļō▒(2013)ņØĆ ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ļČäļ│ĆņŚÉņä£ ļŗżņ¢æĒĢ£ ļ░®ņäĀĻĘĀņØ┤ ļČäļ”¼ļÉśņŚłĻ│Ā, ĻĘĖ ņżæ Streptomyces spp.ļŖö ņŗØļ¼╝ļ│æņøÉĻĘĀ C. gloeosporioides, F. oxysporumņŚÉ ļīĆĒĢśņŚ¼ ĻĘĀņé¼ņāØņן ņ¢ĄņĀ£ļź╝ ļ│┤ņŚ¼ Ļ░ĢĒĢ£ ĒĢŁĻĘĀĒÖ£ņä▒ņØä ļ│┤ņśĆņ£╝ļ®░, ļśÉĒĢ£ ņ╣┤ņĀ£ņØĖ, ņģĆ ļŻ░ļĪ£ņśżņŖż, ĒéżĒŗ┤ņØś ļČäĒĢ┤ļŖźņŚÉ ļīĆĒĢ£ Ļ░ĢĒĢ£ ĒÖ£ņä▒ņØä ļ│┤ņŚ¼ ņ£ĀņÜ®ĒĢ£ ņāØļ¼╝ ņåīņ×¼ņØś ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ļ│┤ņśĆļŗż. ļśÉĒĢ£, Nam ļō▒(2016)ņØĆ ņןņłśĒÆŹļÄģņØ┤ 3ļĀ╣ ņ£ĀņČ®ņØś ļČäļ│ĆņŚÉņä£ ļČäļ”¼ĒĢ£ B. amyloliquefaciens KB3ļŖö lipo-cyclopeptideĻ│äņØś ĒĢŁņāØļ¼╝ņ¦łņØĖ iturin AņÖĆ surfactinļź╝ ņāØņä▒ĒĢśļ®░, ļŗżņ¢æĒĢ£ ņŗØļ¼╝ ĻĘĀļźśļ│æņŚÉ ĻĘĀņé¼ņāØņןņ¢ĄņĀ£ļź╝ ļ│┤ņŚ¼ ļäōņØĆ ņŖżĒÄÖĒŖĖļ¤╝ņØä ļ│┤ņśĆņ£╝ļ®░, ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æņŚÉ ļīĆĒĢśņŚ¼ ļ»ĖņāØļ¼╝ņĀ£ĒÆł Bacillus subtilis QST-713Ļ│╝ ļ╣äĻĄÉĒĢśņŚ¼ ņÜ░ņłśĒĢ£ ļ░®ņĀ£ĒÜ©Ļ│╝ļź╝ ļéśĒāĆļé┤ņ¢┤ ņāØļ¼╝ņĀü ļ░®ņĀ£ņĀ£ļĪ£ ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ņĀ£ņŗ£ĒĢśņśĆļŗż. ļśÉĒĢ£, SwainĻ│╝ Ray (2008, 2009)ļŖö ņåī ļČäļ│ĆņŚÉņä£ ļČäļ”¼ĒĢ£ B. subtilis ĻĘĀņŻ╝Ļ░Ć ļŗżņ¢æĒĢ£ ņŗØļ¼╝ļ│æņøÉĻĘĀņŚÉ ĒĢŁĻĘĀĒÖ£ņä▒Ļ│╝ ņÜ░ņłśĒĢ£ ņØĖņé░Ļ░ĆņÜ®ĒÖö ĒÖ£ņä▒ ļ░Å ĒÜ©ņåīĒÖ£ņä▒ņØä ļ│┤ņØ┤ļ®░ ņ×æļ¼╝ņØś ņāØņןņØä ņ┤ēņ¦äĒĢ£ļŗżĻ│Ā ļ│┤Ļ│ĀĒĢśņśĆļŗż. ņØ┤ņÖĆ Ļ░ÖņØ┤ Ļ│żņČ® ņןļé┤ ļ»ĖņāØļ¼╝ņØś ņÜ░ņłśņä▒ņŚÉ ļīĆĒĢ┤ ļŗżņ¢æĒĢ£ ņé¼ļĪĆņŚÉ ņØśĒĢ┤ Ļ▓Ćņ”ØļÉśņŚłļŗż. ļö░ļØ╝ņä£ ļ│Ė ņŚ░ĻĄ¼ļŖö ņāØļ¼╝ņĀü ļ░®ņĀ£ņĀ£ņØś ņāłļĪ£ņÜ┤ ĒāÉņāēņØś Ļ░ĆļŖźņä▒ņØä ņĀ£ņŗ£ĒĢśņśĆĻ│Ā, ĒÖöĒĢÖļåŹņĢĮĻ│╝ ļ╣äļŻīļź╝ ļīĆņŗĀĒĢĀ ņłś ņ׳ļŖö ņāØļ¼╝ļ╣äļŻīņØś Ļ░ĆļŖźņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż
ņÜö ņĢĮ
Ļ│żņČ®ņØĆ ņןļé┤ņŚÉ ņä£ņŗØĒĢśĻ│Ā ņ׳ļŖö ļ»ĖņāØļ¼╝Ļ│╝ ņāüĒśĖņ×æņÜ®ņØä ĒåĄĒĢ┤ Ļ│ĄņāØĒĢśļŖö Ļ▓āņ£╝ļĪ£ ņĢīļĀżņĀĖ ņ׳ņ£╝ļ®░, ņØ┤ļ¤¼ĒĢ£ Ļ│ĄņāØņ×ÉļŖö Ļ│Ąņ¦äĒÖöļź╝ ĒåĄĒĢśņŚ¼ ĻĘ╣ĒĢ£ ĒÖśĻ▓ĮņŚÉņä£ļÅä ņä£ņŗØņØä Ļ░ĆļŖźĒĢśĻ▓ī ĒĢ£ļŗż. ņØ┤ļ¤¼ĒĢ£ Ļ┤ĆņĀÉņŚÉņä£ ĒåĀņ¢æ ņåŹņŚÉņä£ ļČĆņŚĮĒåĀņÖĆ ņŗØļ¼╝ ņ×öņ×¼ļź╝ ļ©╣Ļ│Ā ņé¼ļŖö ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ņןļé┤ņŚÉ ņĪ┤ņ×¼ĒĢśļŖö Ļ│ĄņāØņ×ÉļŖö ņŗØļ¼╝ļ│æņøÉĻĘĀņØä ļ░®ņĀ£ĒĢśļŖö ļŹ░ ņ£ĀņÜ®ĒĢ£ ļ»ĖņāØļ¼╝ņØ┤ ņĪ┤ņ×¼ĒĢĀ Ļ▓āņ£╝ļĪ£ ņāØĻ░üļÉ£ļŗż. ļö░ļØ╝ņä£, ņŗØļ¼╝ļ│æņøÉĻĘĀņŚÉ ļīĆĒĢ┤ ĒÖ£ņä▒ņØä Ļ░¢ļŖö ņ£ĀņÜ® ļ»ĖņāØļ¼╝ 10ņóģņØä ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ņåīĒÖöĻĖ░Ļ┤Ć ņĀäņן, ņżæņן, Ēøäņןņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ĒĢśņśĆļŗż. ļČäļ”¼ļÉ£ 10ņóģņØś ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØĆ ņ£Āļ¼ś Ļ▓ĆņĀĢņØä ĒåĄĒĢśņŚ¼ ĒåĀļ¦łĒåĀ ņ×┐ļ╣øĻ│░ĒīĪņØ┤ļ│æ, ļ░░ņČö ļ┐īļ”¼Ēś╣ļ│æ, Ļ│ĀņČö ĒāäņĀĆļ│æ, Ļ│ĀņČö ņŚŁļ│æņŚÉ ļīĆĒĢśņŚ¼ Ļ░ĢļĀźĒĢ£ ĒĢŁĻĘĀ ĒÖ£ņä▒ņØä ĒÖĢņØĖĒĢśņśĆļŗż. 10ņóģņØś ĒĢŁĻĘĀĒÖ£ņä▒ ļ»ĖņāØļ¼╝ņØĆ ĒśĢĒā£ņĀü ĒŖ╣ņä▒Ļ│╝ 16s rRNA gene ļČäņäØņ£╝ļĪ£ BacillusņåŹ 4ņóģ, PaenibacillusņåŹ 3ņóģ ļ░Å StreptomycesņåŹ 3ņóģņ£╝ļĪ£ ļÅÖņĀĢļÉśņŚłļŗż. ņ£ĀņÜ® ļ»ĖņāØļ¼╝ņØĆ ņØĖņé░ Ļ░ĆņÜ®ĒÖö, indole-3-acetic acid, siderophore ņāØņä▒ ĒÖ£ņä▒ņØ┤ ņÜ░ņłśĒĢśļ®░ ņ¦äĻĘĀ ņÖĖļ¦ēĻ░ĆņłśļČäĒĢ┤ ĒÜ©ņåīņØĖ ╬▓-1,3-glucanase, pretease ĒÖ£ņä▒ņØä ļ│┤ņśĆļŗż. 10ņóģņØś ņ£ĀņÜ® ļ»ĖņāØļ¼╝ ņżæ, DM152 ĻĘĀņŻ╝ļŖö ĒåĀļ¦łĒåĀņÖĆ Ļ│ĀņČö ņŗØļ¼╝ņ▓┤ņØś ļ¬©ļōĀ ĻĖ░Ļ┤ĆņŚÉņä£ ņāØņןņØä ņ┤ēņ¦äņŗ£ņ╝░ļŗż. ļö░ļØ╝ņä£, ņןņłśĒÆŹļÄģņØ┤ ņ£ĀņČ®ņØś ņåīĒÖöĻĖ░Ļ┤Ćņ£╝ļĪ£ļČĆĒä░ ļČäļ”¼ļÉ£ 10ņóģņØś ņןļé┤ ļ»ĖņāØļ¼╝ņØĆ ņāØļ¼╝ĒĢÖņĀü ļ░®ņĀ£ņĀ£ ļ░Å ņāØļ¼╝ļ╣äļŻīņØś ĒÖ£ņÜ® Ļ░ĆļŖźņä▒ņØä ļéśĒāĆļé┤ņŚłļŗż.

 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






