


서론
유자에 발생하는 주요 병해는 검은점무늬병, 더뎅이병, 역병, 잿빛곰팡이병 등이 있다(Yu 등, 1993). 특히 검은점무늬병과 더뎅이병은 잎, 가지뿐만 아니라 과실에 발병하여 수량과 상품성을 떨어뜨림으로써 경제적 피해가 심각하다. 검은점무늬병 병원균은 Diaporthe citri이며, 대부분의 감귤류를 가해한다. 유자 과실에 나타나는 검은점무늬병 병징은 점들로 보이는 흑점형, 딱지 모양의 니괴형, 흘러내리는 누반형 등 세 가지가 있다(Gopal 등, 2014; Kwon 등, 2003). 이 병은 15년 이상의 노령 유자나무 마른 가지에서 주로 발생하며, 죽은 가지에서 형성된 병자각 속의 병포자가 2차 전염원이 된다(Hur와 Park, 2005a; Park 등, 2014). 병포자 분산은 비바람이 원인인데, 6월 중순부터 8 월 하순 사이의 강우, 소나기, 관수가 중요하며, 특히 강수량과 상대습도가 포자의 분산과 발병을 좌우한다(Hur와 Park, 2005b; Kwon 등, 2003; Park 등, 2014).
습윤은 포자 발아, 부착기 형성 및 발병에 중요한 환경요인이며, 특히 발병 경보 시스템에서 감염을 결정한다(Arauz와 Sutton, 1990). 반면 건조는 식물병원균 포자 발아를 억제하고 발병을 감소시킨다(Kim 등, 2015; Kuhara, 1999; Mondal 등, 2007). Koizumi (1980)가 개발한 유자 검은점무늬병 예측모델인 MELAN은 포자 비산량을 추정한 후, 입력한 온도 및 습윤 지속기간을 근거로 발아율을 계산하여 최종적으로 발병을 예측한다. 이 모델에 따르면 포자 발아율 10%를 달성하지 못하는 온도와 습윤 조건에서 검은점무늬병은 거의 발병하지 않지만, 발아율 50% 이상을 달성하는 온도와 습윤 조건에서는 발병이 확실하다. 또한 병자각에서 누출된 유자 검은점무늬병원균 병포자 발아율은 건조가 지속됨에 따라 점차 감소하는데(Kuramoto와 Yamada, 1975), 기주 표면에 부착한 병원균 포자는 맑은 날에는 4시간, 흐린 날에는 24시간 이상 건조가 지속되면 발아가 억제된다는 결과(Koizumi, 1980)를 근거로 건조가 지속되는 기주 표면에서 포자 발아율은 시간당 20%씩 감소한다는 논리가 MELAN에 포함되었다.
유자 포장에서 6-8월 중 병자각에서 누출된 병원균 포자는 습윤에서 건조로 급변하는 환경에 처할 수 있다. 예컨데 포장에서 밤새 젖었던 잎이 낮에는 말라 습윤-건조 싸이클이 반복되거나, 간헐적인 소나기 이후 급작스런 건조로 인해 습윤이 중단된다. 발아 과정 중 습윤중단은 분생포자 발아율 및 발아관 신장을 저해하고, 사과 붉은곰팡이병과 밀 녹병 발병을 억제한다(Kim 등, 2015). 포자가 건조에 견디는 능력은 병원균 종류에 따라 매우 다양한데, 건조에 민감한(xero-sensitive) 곰팡이 중 대표적인 사과 겹무늬썩음 병원균 Botryosphaeria dothidea은 한 시간 습윤 중단만으로도 포자 발아율 및 발아관 길이가 현저히 감소했다(Kim 등, 2015). 반면, 건조에 저항성(xero-resistant) 병원균인 Alternaria pori는 매일 20시간 건조가 이틀 연속되더라도 포자 발아율은 60% 이상을 유지하였다(Bashi와 Rotem, 1974).
남해안 일대의 우리나라 유자 생산 현장에서 사용가능한 검은점무늬병 방제용 친환경 제재로는 석회유황합제, 기계유 유제, 보르도액, 동제 등이 있다. 다른 친환경 대안으로는 저농약 방제가 있는데, 감염 경보가 발령될 때만 치료용 살균제로 방제하는 것이다. 감염 예측은 유자 포장에서 실시간 측정한 온도 및 수분존재시간으로 예측모델을 구동시켜 모델이 즉각 감염을 판단하는 것이다. 감염 경보시 모델은 약제 살포를 권고하는데, 경보 후 1-2일 이내에 치료용 약제를 살포함으로써 이미 발아를 시작한 포자를 포함하여 병원균을 방제하는 것이 적기방제이다. 일본에서는 유자 검은점무늬병 감염 예측에 MELAN을 활용 중이지만(Kuhara, 1999), 우리나라는 적기방제를 위한 예측 모델을 활용하지 않고 있다.
따라서, 본 연구는 저농약 유자 생산을 위한 검은점무늬병 적기방제 방안으로서 일본에서 개발한 MELAN을 우리나라 유자 생산 현장에 적용하고자 국내 균주를 사용하여 포장에서 나타나는 다양한 건조, 습윤 중단, 온도의 변화에 따른 포자 발아율로 유자 검은점무늬병 감염을 판단하고자 하였다. 병자각으로부터 누출 직후 다양한 건조에 처한 포자가 언제까지 발아력을 유지하는지, 습윤-건조 싸이클에서 습윤중단이 포자 발아에 얼마나 영향이 미치는지를 알아보고자 한다. 이를 위해 실내에서 건조 및 습윤중단 후 검은점무늬병 병원균 포자 발아율을 측정하고, 다양한 온도 및 수분존재시간에서 포자 발아율 10%와 50%에 도달하는데 필요한 국내 균주의 적온과 최소 수분존재시간을 구명하고자 한다. 본 연구 결과는 남해안에서 발병하는 유자 검은점무늬병 감염을 판단하는 우리나라에 적용 가능한 발병 모델 작성에 필요한 기초 자료가 될 것이다.
재료 및 방법
병원균 배양
Diaporthe citri는 2015년 11월 전남 고흥군 풍양면 고흥 농업기술센터 인접 유자 과원(34°34’02.6’’N 127°15’33.7”E)에서 검은점무늬병 병징이 확실한 유자나무 가지로부터 분리하였다. 병원균 순수분리를 위해 100 μg/g streptomycin을 첨가한 감자한천(PDA) 배지에 배양하였다. 병원균 포자 생성을 위해 12시간/일 주기로 배양기에 자외선을 조사하여 25°C에서 2주간 배양 후, 병자각으로부터 노랗게 누출된 포자퇴로부터 병포자를 수확하여 2×104 conidia/ml 농도로 포자현탁액을 제조하여 실험에 사용하였다.
건조 시간에 따른 병포자 발아율 측정
병원균 포자가 병자각에서 누출 직후 건조에 처한 상황을 가정하여 건조기간에 따른 포자 발아율을 측정하였다. 준비한 포자현탁액을 슬라이드글라스 세 부위에 각 20 μl씩 분주 후, 클린 벤치에서 15분간 방치하여 현탁액을 말렸다. 건조 처리는 상대습도 45-55%로 유지되는 25°C 배양기에 0, 3, 6, 9, 12, 15, 24, 48시간 동안 두었다. 슬라이드글라스 위의 건조 처리 포자들의 발아율 확인을 위해, 각 시간별로 건조 처리가 완료된 슬라이드를 젖은 페이퍼 타올이 깔린 밀봉된 플라스틱 상자에 넣고, 25°C 배양기에서 24시간 동안 발아시켰다. 밀봉된 플라스틱 상자 내부는 상대습도 90-95%를 유지함으로써 처리된 포자들이 발아하기에 충분한 환경이었다. 24시간 습윤까지 모든 처리가 완료된 포자들은 광학 현미경(Nikon H600L, Nikon, Japan)에서 100개 포자 당 발아율을 측정하였다. 실험은 같은 방법으로 총 3회 반복하였다.
10시간 습윤 처리 후 습윤중단(건조) 시간에 따른 병포자 발아율 측정
병원균 포자가 습윤 환경에서 건조 환경으로 급변하는 상황을 모의하여 다양한 습윤중단(건조) 시간 처리에 따라 포자 발아율을 측정하였다. 우선, D. citri 포자현탁액을 슬라이드글라스 세 부위에 분주한 후 일괄 10시간 동안 습윤 처리하였는데, 습윤 처리는 젖은 페이퍼 타올을 깔아 내부 습도가 90-95%로 유지되는 밀봉 플라스틱 상자에 슬라이드글라스를 10 시간 동안 두었다. 습윤중단(건조) 처리는 습윤 처리가 완료된 슬라이드글라스를 플라스틱 상자에서 꺼내어 25°C 배양기에 두었는데, 건습구 온도계로 수시 확인하여 배양기 내부 상대습도를 45-55%로 유지하였다. 습윤중단(건조) 처리 기간은 각각 0, 1, 2, 3, 4, 5, 6시간이었다. 습윤중단 처리가 완료된 포자들의 발아율 확인을 위해 90-95% 상대습도가 유지되는 플라스틱상자에 넣어 25°C에서 24시간 동안 배양한 후 현미경으로 검경하였다. 실험은 총 3 회 반복하였다.
10% 및 50% 발아율 달성을 위한 다양한 온도와 습윤 존재시간 실험
Koizumi (1980)에 따르면, 검은점무늬병 예측 모델인 MELAN의 발병 위험 기준은 포자 발아율에 따라 결정된다. 즉, 10% 포자 발아율은 감염에 필요한 최소 수준이며, 50% 포자 발아율은 발병이 확실한 수준이다. 따라서 10% 또는 50% 포자 발아율 달성에 필요한 최소 수분존재시간을 15°C, 20°C, 25°C, 30°C, 35°C에서 확인하였다. 병원균 포자 현탁액을 각 플레이트당 100 μl씩 유자 과피배지에 도말한 후 3, 6, 12, 15, 18, 24 시간에 도달했을 때 포자 발아율을 측정하였다. 유자 과피배지는 과피 400 g에 멸균수 100 ml을 첨가하여 분쇄한 과피 추출액 100 ml과 멸균수 400 ml 그리고 한천 파우더(Difco Laboratories, Detroit, MI, USA) 7.5 g을 혼합하여 제조하였다. 포자현탁액을 접종한 유자 과피배지를 각 온도별로 조절된 인큐베이터에 정해진 시간 동안 배양한 후, 배지를 1 cm × 1 cm 크기로 잘라 슬라이드글라스에 올려 300개 포자 중 발아한 포자수를 현미경 아래서 계수하여 온도 및 시간 별로 포자 발아율을 측정하였다. 실험은 총 3 회 반복 실시하였다. 실험에서 측정한 각 온도별로 병포자가 10%와 50% 발아에 도달하는데 필요한 최소 수분존재시간을 각각 X (온도) 축과 Y (최소 수분존재시간) 축으로 하는 이변량 변수(x, y)로 만들어 이를 점도표로 표현하였다. 작성된 점도표로부터 다양한 온도에 따라 포자 발아 10%와 50%에 도달하는데 필요한 최소 수분존재시간을 추정하는 회귀식을 (Microsoft Excel 2007, Microsoft Corp. Redmond, WA, USA)로 작성하였다.
결과
병자각에서 분생포자가 누출된 직후, 건조 시간 증가에 따른 포자 발아율을 모의하는 실내실험 결과를 Fig. 1에 정리하였다. 건조 처리를 하지 않은(0-24시간) 처리구에서 포자 발아율은 56.6%이었던 반면, 3시간부터 15시간까지 3시간씩 건조 시간이 길어질수록 발아율은 44.7%에서 5.2%까지 점차 떨어졌다. 특히, 24시간 건조 처리는 4.0%, 48시간 건조 처리는 1%까지 포자 발아율이 감소하여 거의 발아한 포자가 없었다(Fig. 1). 발아율 10% 이하는 발병을 거의 기대할 수 없으므로 15시간 이상 건조는 발병을 초래하지 않을 것으로 추정된다.
Fig. 1
The percent of conidial germination of Diaporthe citri on the various dry periods with 45-55% relative humidity. The dry periods were 0, 3, 6, 9, 12, 15, 24, and 48 h. After the dry treatments, the treated conidia were allowed to germinate with moistures for 24 h to measure the treated spore’s potentials of germination.

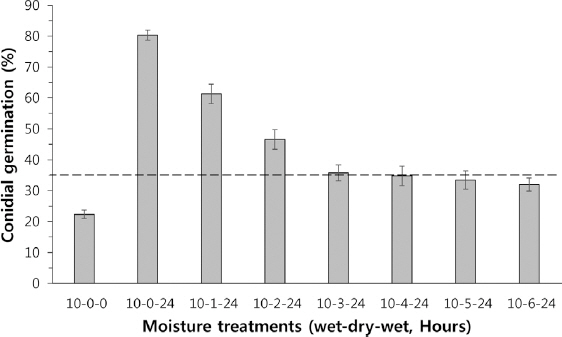
병자각에서 분생포자가 누출 직후 10시간 동안 습윤 환경에 노출되었다가 0-6시간까지 한 시간 간격으로 습윤중단(건조) 처리한 포자들의 발아율 결과를 Fig. 2에 나타냈다. 10시간 습윤만 처리(10-0-0, wet-dry-wet)에서 포자 발아율은 22%였고, 이후 24시간 습윤을 연장한 처리(10-0-24)에서는 전체 포자 중 80%가 발아하였다. 10시간 습윤과 24시간 습윤 사이에 40-45% 상대습도로 1, 2, 3시간 각각 습윤중단(건조) 처리한 구간의 발아율은 각각 60% (10-1-24), 47% (10-2-24), 35% (10-3-24)로 습윤중단 시간이 길어질수록 포자 발아율이 점차 떨어졌다. 한편, 습윤중단(건조) 기간이 4-6시간인 처리들(10-4-24, 10-5-24, 10-6-24)에서 포자 발아율은 35%에서 처리가 길어질수록 점차로 떨어졌다(Fig. 2). 초기 10시간 습윤 처리에서 이미 발아한 22% (10-0-0)를 감안한다면, 3시간 이상 습윤중단 처리된 포자들은 이후 충분한 습윤을 주더라도 발아율이 추가로 약 10% 증가된 후 더 이상 발아하지 않았다.
Fig. 2
The percent of conidial germination of Diaporthe citri treated the various hours of the moisture interruption. The conidia were treated with wet-dry-wet sequence which is supplied moisture for 10 h, then interrupted moisture for 0, 1, 2, 3, 4, 5, and 6 h with 45-55% relative humidity depending on the treatment, finally allowed to germinate with 24 h of wetness.
발병이 거의 되지 않을 10% 이하 포자 발아에 필요한 온도 및 수분존재시간 조건과 심각한 발병이 예측되는 50% 이상 포자 발아에 필요한 온도 및 수분존재시간의 관계를 알아보았다(Fig. 3). 15°C를 제외한 모든 온도에서 24시간 이후 포자 발아율은 70-80%에 도달하였다. 가장 빠른 발아 속도를 보인 온도는 30°C였는데, Fig. 3의 Y축 포자 발아율 50%을 가로지르는 선과 30°C 선이 13.4시간에 교차하여 포자 발아율 50%에 도달하였다. 한편 15°C에서는 24시간 습윤에서도 최대 14% 포자만이 발아하였다. Fig. 3에 표기된 10%와 50% 포자 발아율 수평 점선과 각 온도별 발아율 실선이 만나는 수분존재시간의 X축 좌표 지점을 각 온도별로 발아율 10% 및 50%를 달성하는데 필요한 최소 수분존재시간으로 추정하였다. 20, 25, 30, 35°C의 온도에서 포자 발아율 50%에 도달하는데 필요한 최소 수분존재시간은 각각 19.5, 15.6, 13.4, 14.9시간이었고, 15, 20, 25, 30, 35°C에서 포자 발아율 10%에 도달하는데 필요한 최소 수분존재시간은 각각 21.8, 10.5, 7.8, 7.6, 8.0시간이었다(Fig. 3). 처리한 15-35°C 온도에서 10% 및 50% 발아율 달성에 필요한 최소 수분존재시간을 (x, y)로 하는 이변량 변수로 만들고, 이를 각각 X축과 Y 축으로 하는 점도표를 Fig. 4의 원형(10% 포자 발아율, 5점) 및 삼각형(50% 포자 발아율, 4점)로 표기하였다. 원형과 삼각형을 통과하는 최단거리의 추세선이 10%와 50% 포자발아율 달성을 위해 다양한 온도 조건에서 필요한 최소 수분존재시간을 추정하는 회귀식이라고 할 수 있다. 포자발아율 10%와 50% 달성을 위한 두 회귀식은 다음과 같다.
Fig. 3
The conidial germination of melanose pathogen, Diaporthe citri, according to the temperature regimes (15, 20, 25, 30, 35C) and wetness periods (3, 6, 9, 12, 15, 18, and 24 h) on yuzu feel agar medium. The lines of 10% and 50% of germination is to find the minimum required wetness for the five temperature regimes to achieve 10% and 50% of the spore germination.

Fig. 4
Regressions of 10% conidial germination (●) and 50% conidial germination (▲) of Diaporthe citri according to wetness and temperature conditions. Dots and triangles are required minimum wetness periods for 10% and 50% germination at 15, 20, 25, 30, 35C, respectively.

여러 온도들에서 Wh10% 회귀선보다 짧은 수분존재시간이 주어진다면, 포자발아율이 10%를 넘지 못할 것이며 병은 발생하지 않으리라 예측되며, Wh50% 회귀선보다 더 긴 수분존재시간이 주어지면 포자발아율은 50%가 넘고 발병할 가능성이 매우 높을 것으로 예측된다. Wh10% 회귀선과 Wh50% 회귀선 사이의 수분존재시간에서는 포자발아율이 10%는 넘고 50%는 달성하지 못할 것이며 발병할 위험이 있는 환경이다.
고찰
건조가 지속될수록 포자 발아율은 점차 감소하였으며, 15시간 이상 건조는 발아율을 10% 이하로 떨어뜨려, 발병하지 않을 것이다. 또한 10시간 습윤 후, 3시간 이상 습윤중단(건조)시키면 포자 발아율은 35% 이하로 떨어져, 발병할 위험이 없진 않으나 어려울 것으로 예측된다. 다양한 온도와 수분존재시간 변동 자료로부터 10% 포자가 발아할 수 있는 가장 짧은 수분존재시간은 4.5시간이며, 이 때 온도는 29.2°C로서 어떤 온도보다도 수분존재시간이 가장 짧았다. 이것이 포자 발아율 10%의 적온과 최소 수분존재시간이다. 같은 방법으로 발병이 확실한 50% 포자 발아율 조건에서 수분존재시간은 13시간이 가장 짧았고 이 때 온도가 29.1°C로서 최적 온도였다. 한편, MELAN에서 일본 D. citri 균주의 포자 발아율 10%의 최적 온도는 25°C였고, 이 온도에서 최소 수분존재시간은 2.7시간이었다. 또한, 50% 포자 발아율의 최적 온도는 25°C였고, 최소 수분존재시간은 4.3시간이었다(Kuramoto와 Yamada, 1975). 우리나라와 일본의 유자 검은점무늬병원균 균주를 비교하면, 국내 균주는 일본 균주보다 적온이 약 4.1-4.2°C 높고, 같은 수준의 발아율을 달성하는데 필요한 최소 수분존재시간은 1.8-8.7시간 더 길어 우리나라 균주가 상대적으로 고온이 적온이고 더 오랜 기간 수분을 요구하였다. 결국 일본 균주를 이용한 MELAN을 유자 검은점무늬병 적기방제를 위해 우리나라에 바로 적용하는 것은 적절치 않으며, 우리 균주를 이용한 유자 검은점무늬병 모델 개발이 필요하다.
본 실험 결과와 다른 식물 병원성 곰팡이 건조 실험 결과를 비교하면 45-55% 상대습도로 12시간 건조 처리했을 때 본 실험에서 D. citri 발아율이 15.3%이었다. 한편, 당근 검은잎마름병인 Alternaria dauci는 약 10%(Park 등, 2011), 핵과류 오갈병인 Monilinia fructicola는 12%로서 D. citri 보다는 약간 더 건조에 민감하였다. 습윤 중단의 경우 사과 겹무늬썩음병원균 Botryosphaeria dothidea은 한 시간 습윤 중단만으로도 포자 발아율이 20%이상 감소하였는데(Kim 등, 2015) 이는 본 실험의 D. citri 결과와 비슷하였다. 따라서 D. citri는 건조에 민감한(xero-sensitive) 곰팡이였다.
본 연구에서 건조는 25°C에서 상대습도 45-55%로 처리하였는데, 이는 병자각이 형성되는 6월 중순부터 8월 하순 사이의 유자 포장은 과수의 증산으로 인해 맑은 날에도 대략 40-60% 상대습도가 유지되므로 이를 모사하였다. 한편, Grindle과 Good (1961)이 맥류 붉은곰팡이 병원균인 Fusarium graminearum 건조 처리를 상대습도 45% 뿐만 아니라 상대습도 0%와 15% 등 다양한 상대습도로 실시하였다. 25°C에서 12시간 동안 상대습도 45%로 건조 처리한 F. graminearum 포자 발아율은 10%에 불과했지만, 상대습도 0%와 15%로 12시간 동안 건조 처리하면 발아율은 각각 80%, 45%로 오히려 증가하였다. 즉 상대습도 15% 또는 그 이하로 건조 처리하면 포자발아율이 떨어지지 않았고 도리어 병원균을 휴면상태로 유도하였다. 따라서 건조 처리시 상대습도를 얼마로 할 것인지는 매우 중요하며, 과실이 자라는 건조한 포장 환경의 습도를 고려하여야 한다.
우리가 수행한 실내실험은 병원균 포자 발아율을 슬라이드글라스와 유자 과피 배지에서 측정한 것이다. 우리는 동일한 병원균을 감귤 과실에 접종하여 발아율을 측정한 바 있으나, 수차례의 반복에도 불구하고 유자 과피 위에서 병원균 발아를 직접 관찰하거나 유자 과실에서 검은점무늬병을 발병시킬 수 없었다. 다양한 온도와 수분존재시간에 따른 검은점무늬병 감염 혹은 발병을 판단하려면 실내 실험에서 병원균 발아, 감염, 발병 등 일련의 단계를 구현하는 것이 바람직하나, 본 실험에서는 인위 접종 후 유자 과실에서 발아와 발병을 관찰할 수 없어 유자 과실대신 유자 과피를 갈아 배지를 만들고 그 위에서 MELAN 모델이 제시하는 감염 판단 기준인 10% 및 50% 포자발아율에 도달하는 온도와 수분존재시간으로 결과를 도출하였다.
유자 검은점무늬병은 어린 나무보다는 수령 15 년 이상의 유자 나무에서 더 빈번하게 발생하는데, 고령목에서 생산된 유자가 향과 품질이 뛰어나므로 고령목 유자의 저농약 친환경 재배는 앞으로 반드시 달성해야 할 도전 과제이다. 아직까지는 우리나라에서 유자 검은점무늬병 저농약을 실현하기 위한 적기방제를 하지 않고 있다. 본 연구는 10%와 50% 포자 발아에 필요한 다양한 온도와 수분존재시간 결과를 통해 검은점무늬병의 감염 시작 및 심각한 감염을 판단하는 기준을 제안하였다. 앞서 언급한 것처럼 우리나라 남부지방에서 분리한 국내 균주가 일본 균주에 비해 적온이 높고 건조에 더 민감하며 같은 발아율 달성에 더 오랜 기간의 수분존재시간이 필요하였다. 더불어서 전남지역 포장에서 수행된 죽은 유자 나뭇가지에서 병포자 형성에 대한 연구(Hur와 Park, 2005a)와 D. citri 포자의 비산 및 병자각 형성 연구(Hur와 Park, 2005b) 결과는 한국형 모델을 만드는데 필요한 매일 죽은 가지가 새롭게 생기는 양을 측정하기 위한 계산과 병원균 포자 비산량 계산에 활용할 수 있다. 결국 남부지방 검은점무늬병 감염을 판단하기 위한 한국형 예측 모델을 만들려면, 본 연구의 실내 발아실험 결과뿐만 아니라 Hur와 Park (2005a, 2005b)의 포장 연구 결과를 포함하여야 하며, 발병 예측을 통해 궁극적으로 유자 검은점무늬병 저농약 방제를 실현할 수 있을 것이다.
요약
유자 검은점무늬병은 Diaporthe citri가 원인균이며, 과실에 발병하여 심각한 피해를 초래한다. 병자각으로부터 분생포자가 누출될 때, 습윤은 병원균 감염에 중요한 환경 요소이다. 발아과정 중 병포자가 건조에 처하거나 습윤중단이 발생할 때 발아율에 어떤 영향을 주는지 알아보았다. 건조 처리는 포자현탁액을 슬라이드 글라스에 0-48시간 동안 처리하였다. 습윤 중단 처리는 포자현탁액을 습윤에 10시간 먼저 노출한 후, 0-6 시간 건조시켜 발아과정에서 습윤을 중단시켰다. 15시간 이상 건조 처리한 포자 발아율은 10% 이하였다. 3시간 이상 습윤 중단 처리된 포자 발아율은 35% 이하였다. 다른 식물병원성 곰팡이와 비교하면, D. citri는 건조에 민감한 곰팡이다. 유자 과피 배지 위에 다양한 온도(15, 20, 25, 30, 35°C)별로 습윤 존재시간(3, 6, 9, 12, 15, 18, 24 hr)을 다양하게 처리하여 포자발아율을 측정하였다. 발아율 10%와 50%를 달성하기 위한 최소 수분존재시간은 각각 4.5시간과 13시간이며, 이 때 적온은 각각 29.2°C와 29.1°C였다. 다양한 온도에서 발아율 10%와 발아율 50% 달성에 요구되는 최소 수분존재시간을 결정하는 추정식은 각각 Wh10% = 0.082 × T2 - 4.8025 × T + 74.861과 Wh50% = 0.1093 × T2 - 6.3762 × T + 106.08이었다.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print






